Click through the PLOS taxonomy to find articles in your field.
For more information about PLOS Subject Areas, click here .
Loading metrics
Open Access
Peer-reviewed
Research Article

Contextual Interference in Complex Bimanual Skill Learning Leads to Better Skill Persistence
* E-mail: [email protected]
Affiliation KU Leuven, Movement Control and Neuroplasticity Research Group, Department of Kinesiology, Group Biomedical Sciences, Leuven, Belgium
Affiliations KU Leuven, Movement Control and Neuroplasticity Research Group, Department of Kinesiology, Group Biomedical Sciences, Leuven, Belgium, KU Leuven, Leuven Research Institute for Neuroscience & Disease (LIND), Leuven, Belgium
- Lisa Pauwels,
- Stephan P. Swinnen,
- Iseult A. M. Beets
- Published: June 24, 2014
- https://doi.org/10.1371/journal.pone.0100906
- Reader Comments
The contextual interference (CI) effect is a robust phenomenon in the (motor) skill learning literature. However, CI has yielded mixed results in complex task learning. The current study addressed whether the CI effect is generalizable to bimanual skill learning, with a focus on the temporal evolution of memory processes. In contrast to previous studies, an extensive training schedule, distributed across multiple days of practice, was provided. Participants practiced three frequency ratios across three practice days following either a blocked or random practice schedule. During the acquisition phase, better overall performance for the blocked practice group was observed, but this difference diminished as practice progressed. At immediate and delayed retention, the random practice group outperformed the blocked practice group, except for the most difficult frequency ratio. Our main finding is that the random practice group showed superior performance persistence over a one week time interval in all three frequency ratios compared to the blocked practice group. This study contributes to our understanding of learning, consolidation and memory of complex motor skills, which helps optimizing training protocols in future studies and rehabilitation settings.
Citation: Pauwels L, Swinnen SP, Beets IAM (2014) Contextual Interference in Complex Bimanual Skill Learning Leads to Better Skill Persistence. PLoS ONE 9(6): e100906. https://doi.org/10.1371/journal.pone.0100906
Editor: Edwin Robertson, Harvard Medical School, United States of America
Received: December 16, 2013; Accepted: May 30, 2014; Published: June 24, 2014
Copyright: © 2014 Pauwels et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was supported by a grant from the Research Program of the Research Foundation – Flanders (Fonds Wetenschappelijk Onderzoek – FWO; http://www.fwo.be/ ) (G0483.10, G0721.12), from the Research Fund of KU Leuven, Belgium (OT/11/071), and Grant P7/11 from the Interuniversity Attraction Poles program of the Belgian federal government. LP is funded by an aspirant fellowship of the Research Foundation – Flanders (FWO). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
Introduction
In the field of practice organization for motor skill learning, contextual interference (CI) has been one of the most frequently discussed topics over the past three decades. The term, CI, was introduced by Battig [1] and refers to the interference that results from performing various tasks or skills within the context of practice. Shea and Morgan [2] were the first to establish this effect in motor skill learning. Since then, numerous investigations led to the finding that introducing high amounts of CI, by means of presenting multiple task variants in a randomized order, leads to inferior performance during acquisition but benefits learning, reflected by better retention and transfer tests, in contrast to a blocked practice schedule [3] , [4] . This phenomenon denotes a distinction between performance and learning and is often referred to as the paradoxical opposing effects of CI during acquisition and retention.
The CI effect is quite a robust phenomenon in laboratory experiments using multi-segment movement tasks, coincident anticipation timing tasks, aiming tasks, movement patterning tasks and tracking tasks [4] , [5] . Several theoretical explanations underlying the CI effect have been proposed of which the elaboration hypothesis [2] , [6] and the action-plan reconstruction hypothesis [7] , [8] are the most prominent ones. The elaboration hypothesis [2] states that the benefits of high CI are due to more elaborative and distinctive processing because multiple tasks reside together in working memory, whereas when practicing under low CI, only one task is present in working memory. However, Lee and Magill [7] proposed the action-plan reconstruction hypothesis stating that action plans will be forgotten because of alternating trials following a random practice schedule. Thus, high amounts of CI will result in more effortful reconstructive processing whereas the action plan in blocked practice will be remembered.
Nevertheless, these two hypotheses may have a common factor namely enhanced cognitive effort and processing when practicing under high CI and decreased cognitive activity and processing when CI is low [9] . Recently, Kantak and Winstein [10] suggested a novel point of view to clarify the distinction between performance and learning by proposing a motor behavior-memory framework that shows the evolution of motor memory processes. The behavior-memory framework highlights the importance of the temporal evolution of memory processes (i.e. encoding, consolidation and retrieval) and states that the efficiency of these processes can be reflected in performance measures at different time points. Such a framework is important because motor memory processes, for example encoding [11] , [12] and consolidation [13] , [14] processes, can be differentially affected by different practice structures.
Although numerous studies have confirmed the CI effect in rather simple tasks, the CI effect is much less explored in complex tasks that require more cognitive effort. Multiple studies have been carried out, supporting either of both hypotheses [4] . However, it remains to be seen which of the two is favorable. In addition, none of these accounts make distinct predictions with respect to the CI effect in more complex tasks [5] . As stated by Wulf and Shea [5] , the key question is whether principles derived from simple task studies can be extended to complex skill learning, which is essential in real life. According to the aforementioned hypotheses, enhanced cognitive activity and processing through high CI are assumed to be critical for enhanced learning. But what if the nature of the task itself is more difficult requiring high attention and memory demands? Would high CI further benefit or rather perturb skill learning? This prompts questions about the boundary conditions of the CI effect. Previous studies examining the effect of CI in complex motor skills have led to contrary results [5] . While some studies succeeded in demonstrating clear benefits of high amounts of CI in complex skill learning [15] – [17] , others did not find any evidence at all [18] , [19] . Both the studies of Albaret and Thon [20] and Tsutsui et al. [21] yielded mixed results. Because these papers are related to our topic, a more in depth discussion about these papers can be found in the discussion section. Yet, Wulf and Shea [5] aimed to get an overall view of the CI effect in tasks of various difficulty levels and noted that random practice is generally effective when learning simple tasks, i.e. with low attention and memory demands, or when a person is experienced in a complex task, requiring reduced memory demands. In line with this notion, Shea et al. [22] proposed that the benefits of CI increased with increasing amounts of practice. This might suggest that a certain level of experience in a complex task is required in order to obtain benefit from randomized practice [22] , [23] .
Since many tasks in daily life require a good coordination between both hands, the purpose of this study was to explore the CI effect in a complex bimanual coordination task. However, defining “complexity” is a tough job [5] . Considering the distribution of brain activations involved in various bimanual tasks, we assume our task to be sufficiently complex as it requires higher cognitive functions [24] . In the current paper, three task variants were divided over multiple days of training. In accordance with both the elaboration and action-reconstruction hypotheses, we hypothesized that practicing under high (i.e. a randomized practice schedule) as compared to low CI will have detrimental effects during acquisition, but will benefit retention performance immediately and seven days after the last practice day. However, as discussed previously, Kantak and Winstein [10] highlighted the importance of the temporal evolution of memory processes. Thus, besides directly comparing retention performance between groups, which encompasses both encoding and consolidation processes, we also looked into processes occurring during the retention interval, i.e. post-acquisition. As one key feature of the CI effect is that temporary performance benefits should be sacrificed for long-term learning benefits [4] , we hypothesized that practicing under a randomized practice schedule will result in better performance persistence across retention intervals. During these post-acquisition processes, a long-term memory representation will be generated and thus, better performance persistence will be an indicator of a more stable memory representation [10] . In view of the previously addressed literature, we assumed this would only be the case when the task is sufficiently practiced. Therefore, we extended the practice schedule beyond those typically used in previous studies. We examined the CI effect in complex skill learning while focusing on the level of skill persistence from the end of acquisition to retention.
Materials and Methods
Forty young, healthy, right-handed subjects (mean age = 19.6±1.3 years; range 18–23 years) took part in the experiment. All subjects were right-handed as determined by the Oldfield Handedness scale [Oldfield, 1971] (mean laterality 85.8±13.6). Subjects were randomly assigned to either of the two groups: blocked practice group (n = 20; 10 female; mean age = 19.4±1 year; mean laterality 85.6±14.7) and randomized practice group (n = 20; 10 female; mean age = 19.8±1.7 year; mean laterality 86.1±12.6) and were blind to the purpose of the experiment. There were no between-group differences with respect to age [ p = 0.357], laterality quotient [ p = 0.924] and gender was equally distributed across groups. Prior to testing, written informed consent was obtained from each subject. The protocol was approved by the local ethical committee of the University of Leuven (KU Leuven), Belgium, and was in accordance with the Declaration of Helsinki (1964).
Instrumentation and task description
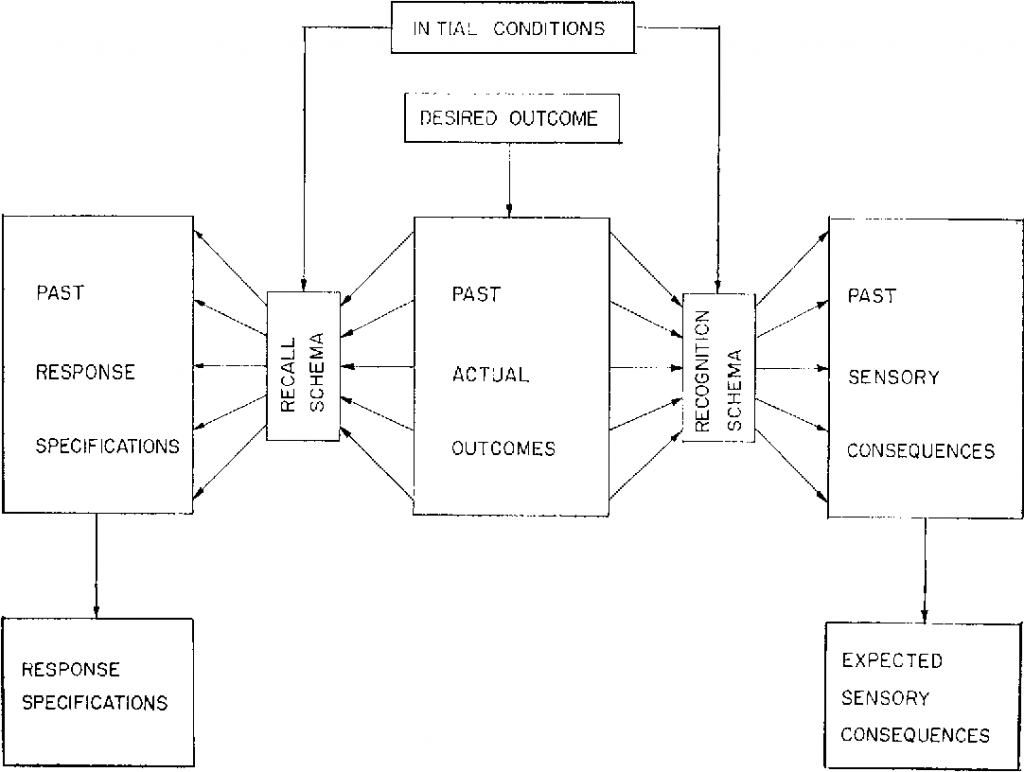
A PC-based visuo-motor bimanual tracking task was used. Subjects were seated in front of a computer screen with both lower arms resting on two custom-made adjustable ramps ( Figure 1A ). The ramps were covered with foam to assure maximal comfort and to minimize fatigue. A dial, consisting of a flat disc (diameter 5 cm) with a vertical peg, was attached at the end of each ramp. The aim of the task was to follow a white target dot along a blue target line on the screen. To perform the required movement, subjects rotated both dials simultaneously by holding each peg between the thumb and index finger. Direct vision of hands and forearms was prevented by placing a horizontal table-top bench over the forearms of the subject. High precision shaft encoders were aligned with the axis of rotation of the dials to record angular displacement (Avago Technologies, sampling frequency = 100 Hz, accuracy = 0.089°). A red cursor showed the current position so that the deviation from the target dot could be corrected. The left dial controlled the vertical component of the red cursor, such that when turning it clockwise, the cursor moved up and when turning it counterclockwise, the cursor moved down. The right dial controlled the horizontal component of the red cursor, such that when turning the dial clockwise or counterclockwise, the cursor moved right and left, respectively. The gain was set to 10 units per rotation, so that 16 complete rotations of both hands were required to complete the target line that consisted of 160 arbitrary units.
- PPT PowerPoint slide
- PNG larger image
- TIFF original image
(A) Task set-up . Subjects were seated in front of a computer screen on which the task was displayed. The response apparatus consisted of two dials which were fixated on a ramp. Direct vision of the forearms was prevented by a horizontal table-top bench. (B) Frequency ratios and coordination directions . Schematic drawing of the target lines shown on the screen, from which subjects can deduct the three frequency ratios (1∶1, 2∶3 and 1∶2) and coordination directions [clockwise (CW) and counterclockwise (CCW)]. The coordination directions inwards (IN) and outwards (OUT) are shown here, but are not a part of the training protocol.
https://doi.org/10.1371/journal.pone.0100906.g001
A blue target line indicated the main coordination directions: both hands could rotate both clockwise (CW), both counterclockwise (CCW), inwards (IN) and outwards (OUT) ( Figure 1B ). The latter two coordination directions were not used in the current training protocol; however, they were used for instruction prior to testing in order to maximize understanding of the rules of the task (see below). Each coordination direction could be performed at different frequency ratios, which was visualized by the slope of the target line ( Figure 1B ). A target line with a 45° slope indicated a 1∶1 frequency ratio, whereby both hands were required to rotate at equal speeds. We used the convention of referring to the left hand first and the right hand second, i.e. L:R. For example, a 1∶2 frequency ratio required the right hand to move twice as fast as the left hand.
Three types of feedback conditions were used: concurrent visual feedback (cFB), after-trial feedback (atFB) and no feedback (NFB) ( Figure 2 ). In all conditions, the blue target line and the white target dot were presented. In each trial, the white target dot was first covered by a yellow cue which indicated whether cFB would be given in the upcoming trial. The cue and target dot remained motionless in the center of the screen for 2 s. No movement was required, but the subject was instructed to plan the movement (planning phase). Then, an auditory cue was provided to indicate the start of the execution phase. The execution phase lasted 9 s. During the execution phase, the white target dot moved with constant speed starting from the center of the display, along the blue target line, towards the periphery. The goal of each trial was to generate the correct direction and speed by turning the dials in order to stay as close as possible to the white target dot. The inter-trial interval (ITI) lasted 3 s in which a black screen was presented. In the cFB condition, current performance was visualized online by a red cursor which contained the most recent information of the subjects' movement track (1 s), upon which movements could be corrected. During atFB trials, no red cursor was shown, but the same blue target line and the white target dot were presented. After-trial FB was provided after each trial by presenting a motionless representation of 1 s consisting of the produced red line, representing the produced movement next to the required target line, indicating the discrepancy between the produced and the required movement. In NFB trials, the blue target line and the moving white target dot were also presented, but neither cFB nor atFB was provided. Thus, in both the atFB and NFB conditions, subjects were required to track the target pathway of the frequency ratio without the guidance of concurrent visual FB.
Concurrent visual feedback, provided by a red cursor indicative of subjects' current position, was only provided in the concurrent visual feedback (cFB) condition. In the after-trial feedback (atFB) condition, a motionless representation of the produced red line was provided after the execution phase while no feedback was provided during the execution phase. In the no feedback (NFB) condition, no concurrent or after-trial feedback was provided. Every trial started with a planning phase of 2 s where a yellow cue, which indicated whether cFB would be given in the upcoming trial, was presented. During the execution phase, the white target dot moved with constant speed along the blue target line for 9 s. In each condition, the inter-trial interval (ITI), i.e. the time between each trial where no movement was required, lasted 3 s. During ITI, atFB was provided for 1 s in the atFB condition. Instead, a black screen was presented in the cFB and NFB condition.
https://doi.org/10.1371/journal.pone.0100906.g002
Study design
Subjects had to learn 3 different frequency ratios (1∶1, 2∶3 and 1∶2) in 2 coordination directions (CW, CCW), i.e. 6 different trial types, over 3 practice days within one week. The 6 trial types were trained either under a blocked (i.e. low CI) or randomized (i.e. high CI) practice schedule, depending on practice group. Prior to testing, subjects were informed about the basic requirements to perform the task, i.e. knowledge of the different directions and their associated rotations (CW, CCW, IN and OUT). No information was given on how to produce the different frequency ratios. To assess whether every subject understood the basic requirements of the task, a familiarization block consisting of 4 trials, i.e. a 1∶1 frequency ratio in each coordination direction (CW, CCW, IN and OUT), was conducted. For an overview of the training protocol, see Figure 3A .
(A) Training protocol . Baseline performance was assessed without concurrent FB (NFB) on day 1 prior to training. The acquisition phase consisted of 3 training days within one week. Because of the fading feedback schedule, all 3 feedback conditions were present during each day of training. Immediate retention (IR) was conducted 5 min after the end of training day 3 and delayed retention (DR) was conducted 7 days later. Both IR and DR consisted of 2 types of retention schedule, i.e. a blocked (IR-B and DR-B) and a random (IR-R and DR-R) schedule. (B) Blocked and randomized practice schedule . Subjects in the blocked practice group were exposed to one frequency ratio in both clockwise (CW) (blocks 1–3) and counterclockwise (CCW) (blocks 4–6) directions per day. In contrast, subjects in the randomized practice group were exposed to all 6 trial types (which were randomly presented) during each block, i.e. 4 trials per trial type in each block during training. The number of different feedback (cFB, atFB and NFB) trials and the degree of fading feedback within each trial type was identical in both groups. Therefore, concurrent feedback (cFB) in the blocked practice group faded over blocks 1 to 3 after which the fading feedback schedule repeated itself during the next 3 blocks. In contrast, in the randomized practice group, fading feedback was distributed over days within each trial type.
https://doi.org/10.1371/journal.pone.0100906.g003
Feedback schedule.
According to the challenge point framework described by Guadagnoli and Lee [25] , providing practice conditions that facilitate performance, for example by presenting high amounts of feedback until the movement representation is relatively stable, will enhance skill learning when a task is more complex. In addition, they state that the processing system is too inefficient at the early stage of learning [26] , and that the learner might not be capable of interpreting information efficiently, leading to more frequent information being needed in order to yield learning [25] . In contrast, with practice (later stage of learning), the ability to process information increases, leading to better prediction to reach the goal [25] , [27] , [28] . Accordingly, high amounts of feedback might then be redundant for the learner [25] , [27] – [29] . Therefore, to prevent reliance on feedback and to optimize learning, we made use of a fading feedback schedule [30] , [31] . That is, for each trial type, we gradually reduced the number of trials in which we provided concurrent visual feedback. There were 72 trials per trial type (i.e. per frequency ratio in either CW or CCW coordination directions), across training for both practice groups. Each practice day consisted of 144 trials with six blocks of 24 trials each ( Figure 3B ). The numbers of cFB trials were gradually reduced (50% for trial 1–24; 33% for trial 25–48 and 21% for trial 49–72) while the NFB trials gradually increased (21%; 33% and 50% respectively) for each trial type ( Figure 3B ). The number of atFB trials was kept relatively constant for each trial type throughout training (29% for trial 1–24, 33% for trial 25–48 and 29% for trial 49–72). In the blocked practice group, feedback faded from block 1 to 3 after which the fading feedback schedule repeated itself for the next trial types, i.e. during every following 3 practice blocks. In the randomized practice group, all 6 trial types were randomly presented during every block across training. That is, during every training day, all 6 trial types were practiced, i.e. each trial type was presented 4 times during each block, and the trial number of each trial type (i.e. 72) was spread across the three training days. The fading feedback schedule in the randomized practice group was therefore spread over training days for each trial type. Concurrent FB was given for 50% of trials on day 1, 33% on day 2, and 21% on day 3 for all trial types. Within each training day, cFB also generally faded, starting with more cFB trials at the beginning of the training day and ending with more NFB trials. The number of cFB, atFB and NFB trials and the degree of fading within each trial type was identical in both groups. In order to see how performance without visual guidance evolved in both practice groups, i.e. where the subjects had to produce movements primarily based on an internal representation of the movement pattern instead of having the opportunity to make online corrections based on external visual information, only trials without concurrent visual feedback (65% of 432 trials), i.e. atFB and NFB, were used for analyses of acquisition phase data. For baseline and retention tests, only NFB trials were presented to subjects in order to prevent learning during these tests from online visual feedback or after trial feedback.
To assess baseline performance, i.e. without prior exercise of the to be trained trial types, subjects had to perform 12 NFB trials, i.e. 2 trials per trial type, in the following blocked order: 1∶1 CW –1∶1 CCW –2∶3 CW –2∶3 CCW –1∶2 CW –1∶2 CCW.
Acquisition phase.
The acquisition phase took 3 training days within one week. Subjects in the blocked practice group learned one frequency ratio per day in the following order: 1∶1 on day 1, 2∶3 on day 2 and 1∶2 on day 3. Each frequency ratio was learned in the CW (blocks 1–3) and in the CCW (blocks 4–6) coordination direction. Subjects in the randomized practice group were exposed to all 6 trial types in a randomized order during every block, i.e. 4 trials per trial type in each block, of each practice day. The number of practice trials for every trial type was equal for both groups. At the end of practice, a total of 432 trials were completed, of which 150 cFB trials and 282 trials without concurrent FB (132 trials atFB and 150 trials NFB). For each trial type, a total of 72 trials were practiced with 25 trials with and 47 trials without concurrent FB (22 trials atFB and 25 trials NFB). Approximately 45 minutes were needed to finish 6 practice blocks. For an overview of the acquisition phase, see figure 3B .
Immediate retention (IR).
Following the acquisition phase at the last day of practice (after a 5 min break), subjects were involved in an immediate retention (IR) test to assess the practiced frequency ratios. Retention accuracy is not only dependent on practice context, but also on the context in which retention is measured, i.e. a blocked or random retention test [2] , [22] . Therefore, 2 retention schedules were used: a blocked IR (IR-B) and a randomized IR (IR-R). Both the IR-B and IR-R consisted of 24 NFB trials, i.e. 4 trials per trial type. During IR-B, the coordination patterns were tested following the same order as during the baseline test. After IR-B (following 1 min of rest), IR-R was conducted in which all learned coordination patterns were presented randomly. IR-B was always tested before IR-R in order to avoid confounding effects for the blocked practice group on a blocked schedule after having contact with a randomized schedule. Both IR-B and IR-R took 6 minutes to complete.
Delayed retention (DR).
A delayed retention (DR) test, which also consisted of a blocked DR (DR-B) and a randomized DR (DR-R), was conducted 7 days after the last day of practice. The two DR tests were exactly the same as the IR tests.
Dependent measures
Data were recorded and analyzed with Labview (8.5) software (National Instruments, Austin, Texas, USA). The x- and y positions of the target dot and the subjects' cursor were sampled at 100 Hz. Offline analysis was carried out using Matlab R2011b and Microsoft Excel 2010. Accuracy was measured by calculating the error rate based on the average target deviation (ATD). For each trial, the target error was measured as the Euclidian distance between the target dot and the cursor position at each point in time and then averaged. Better performance is thus reflected by lower values of ATD. Because our primary focus was on the different frequency ratios and because CW and CCW movements were mainly used to provide an extra dimension of complexity to the task (as subjects needed to alternate between them), we collapsed CW and CCW data within each frequency ratio. Besides, previous research of our lab showed that the difference between CW and CCW coordination directions is negligible [32] . For the acquisition phase analyses, frequency ratio data was averaged across every set of 3 data points in time. This resulted in 16 acquisition phase data points for each frequency ratio (TR1, TR2, …, TR16). As such, TR1 for the 1∶1 frequency ratio consisted of 6 trials (3 CW and 3 CCW) without concurrent FB. Outlier trials (z>2) were discarded (6% in total of which 7% in the blocked practice group and 5% in the random practice group) from the analyses. To reduce the positive skew that was present in our data, data were log-transformed (base 10 logarithm).
Statistical analysis
Frequency ratio..
In order to explore whether time courses of different frequency ratios differed among each other, a full model analysis on all time points was conducted using a 2×21×3 Group (blocked, random) × Time (Baseline, TR1-16, IR-B, IR-R, DR-B and DR-R) × Frequency ratio (1∶1, 2∶3 and 1∶2) repeated measures ANOVA with Group as between-subject factor and Time and Frequency ratio as within-subject factors. First, a Time × Frequency ratio interaction effect [ F (40,1520) = 2.301, p <0.001] was found, indicating that performance over time was different for each frequency ratio. In addition, a Group × Time × Frequency ratio interaction effect [ F (40,1520) = 1.453, p = 0.034] was found indicating that performance over time significantly differed for each frequency ratio per group. This pointed towards a clear difference in time course between the 3 frequency ratios. As such, to investigate between-group differences during the acquisition phase, IR and DR, we decided to conduct separate analyses for these 3 frequency ratios.
In order to assess whether there were group differences prior to practice, baseline performance was analyzed using a 2×3 Group (blocked, random) × Frequency ratio (1∶1, 2∶3 and 1∶2) repeated measures ANOVA.
Acquisition phase data were analyzed using a 2×16 Group (blocked, random) × Time (TR1-16) repeated measures ANOVA. To assess whether learning occurred from baseline to the end of acquisition in both practice groups, a control analysis, a 2×17×3 Group (blocked, random) × Time (Baseline, TR1-16) × Frequency ratio (1∶1, 2∶3 and 1∶2) repeated measures ANOVA, was conducted.
Immediate and delayed retention.
To test whether performance during retention tests differed per practice group, IR and DR were analyzed using a 2×2 Group (blocked, random) × Retention schedule (blocked, random) repeated measures ANOVA.
Planned comparisons.
Finally, we aimed to test whether the random practice group will show more performance persistence during the retention intervals than the blocked practice group. Planned a priori comparisons of least square means were conducted on the full model (2×21×3 ANOVA) to test the hypothesized differential change in performance, i.e. difference in post-acquisition processes between both groups. The two final time points of training, i.e. the end of acquisition (EoA: TR15-TR16), were taken and compared with IR and DR for both practice groups to test the interaction of Group × Time. In order to conduct these partial interaction contrasts, weights were assigned as follows. To assess the factor Group, each practice group was assigned a weight, i.e. 1 for the blocked practice group and -1 for the random practice group. For the repeated measures factor Frequency ratio, contrasts were conducted for each frequency ratio separately leading to a weight of 1 for one frequency ratio and weights of 0 for the remaining two. For the repeated measures factor Time, TR15 and TR16 both received a weight of -1, i.e. EoA, and IR-B (or DR-B for DR) and IR-R (or DR-R for DR) both received a weight of 1 in order to combine these means.
Statistical analyses were conducted using STATISTICA. For all analyses, the probability level was set at p <0.05, 2-sided. When significant effects were found, post hoc analyses were conducted using Tukey HSD.
Performance was compared between the randomized and blocked practice group to test CI in bimanual coordination. Performance differences were tested before practice (baseline), over the course of practice (acquisition phase), and at retention. In addition, planned comparisons were conducted in order to get a view into the efficiency of post-acquisition processes in both practice groups. As the three frequency ratios clearly differed from each other and showed an interaction effect with Group and Time (see methods), they were analyzed and illustrated separately.
The 2×3 Group × Frequency ratio repeated measures ANOVA did not reveal a significant main effect of Group [ F (1,38) = 1.857, p = 0.181], indicating that the performance level at baseline of both groups was comparable (blocked practice group: 1.18±0.19; random practice group: 1.12±0.14). A main effect of Frequency ratio [ F (2,76) = 28.294, p <0.001] was found. Post hoc analyses revealed that the 1∶1 frequency ratio was easier to perform than the 2∶3 frequency ratio ( p <0.001) and 1∶2 was the most difficult frequency ratio prior to practice ( p <0.001 and p = 0.007 compared with the 1∶1 and 2∶3 frequency ratio, respectively), which is in line with previous studies in our research group (e.g. Sisti et al. [32] ). There was no Group × Frequency ratio interaction [ F (2,76) = 0.558, p = 0.575], suggesting that the effect of Frequency ratio was similar for both groups at baseline.
Acquisition phase
For each frequency ratio, the 2×16 Group × Time ANOVA revealed a significant main effect of Time [ F (15,570) = 3.935, p <0.001; F (15,570) = 6.342, p <0.001 and F (15,570) = 5.811, p <0.001 for the 1∶1, 2∶3 and 1∶2 frequency ratio, respectively]. The 2×17×3 Group (blocked, random) × Time (Baseline, TR1-16) × Frequency ratio (1∶1, 2∶3 and 1∶2) control analysis revealed a significant main effect of Time [ F (16,608) = 57.183, p <0.001]. Post hoc analyses indicated performance improvements for each practice group from baseline to TR16 ( p <0.001 for each frequency ratio) ( Figures 4 , 5 and 6 ). For the 1∶1 frequency ratio, there was no significant main effect of Group [ F (1,38) = 0.511, p = 0.479]. However, there was a significant Time × Group interaction effect [ F (15,570) = 3.243, p <0.001]. The randomized group started out with worse performance compared to the blocked practice group but performance became nearly as good as in the blocked practice group after approximately one-third of practice ( Figure 4 ). For the 2∶3 frequency ratio, there was no difference between groups [ F (1,38) = 2.139, p = 0.152]. A trend towards significance was found for the Time × Group interaction [ F (15,570) = 1.635, p = 0.060] in which performance of the randomized practice group showed a less stable pattern along time with higher error peaks during practice (especially on TR5 and TR10, which was the start of training day 2 en 3 respectively for the randomized practice group) compared to the blocked practice group ( Figure 5 ). In the 1∶2 frequency ratio, the overall performance was worse for the randomized compared to the blocked practice group, as indicated by a significant main effect of Group [ F (1,38) = 25.564, p <0.001]. This group difference was more pronounced at the beginning of practice as reflected by the significant Time × Group interaction [ F (15,570) = 3.510, p <0.001], whereby the randomized group showed more improvement than the blocked group ( Figure 6 ).
Error score (ATDlog, i.e. the log-transformed average target deviation) for baseline, acquisition phase (TR1-16), immediate retention (IR) and delayed retention (DR) (mean ± standard error) learned under either a blocked (black circles) or randomized (white squares) practice schedule. Better performance is indicated with lower levels of ATDlog.
https://doi.org/10.1371/journal.pone.0100906.g004
https://doi.org/10.1371/journal.pone.0100906.g005
https://doi.org/10.1371/journal.pone.0100906.g006
Immediate retention
During IR, performance of the randomized practice group was significantly better when performing the 1∶1 (0.76±0.22) and 2∶3 (0.89±0.14) frequency ratio compared to the blocked practice group (0.99±0.14 and 1.03±0.11 for the 1∶1 and 2∶3 ratio, respectively), reflected by a main effect of Group [ F (1,38) = 20.473, p <0.001 and F (1,38) = 13.688, p <0.001 for the 1∶1 and 2∶3 frequency ratio, respectively] ( Figures 4 and 5 , respectively). For the 1∶2 frequency ratio, no difference between groups was found [ F (1,38) = 0.22, p = 0.642] ( Figure 6 ). There was no main effect of Retention schedule [ F (1,38) = 0.292, p = 0.592; F (1,38) = 0.731, p = 0.398 and F (1,38) = 0.296, p = 0.590 for the 1∶1, 2∶3 and 1∶2 frequency ratio, respectively], indicating that randomized versus blocked testing at retention did not affect performance. There was also no interaction effect of Group × Retention schedule [ F (1,38) = 0.077, p = 0.783; F (1,38) = 0.156, p = 0.695 and F (1,38) = 0.029, p = 0.865 for 1∶1, 2∶3 and 1∶2 frequency ratio, respectively].
Delayed retention
Randomized practice resulted in better delayed retention performance of the 1∶1 (0.83±0.18) and 2∶3 (0.95±0.17) frequency ratios compared with the blocked practice group (0.99±0.21 and 1.06±0.12 for the 1∶1 and 2∶3 ratio respectively), reflected by the main effect of Group [ F (1,38) = 8.654, p = 0.006 and F (1,38) = 7.61, p = 0.008 for the 1∶1 and 2∶3 frequency ratio, respectively] ( Figures 4 and 5 ). For the 1∶2 frequency ratio, no between group difference was found [ F (1,38) = 0.089, p = 0.767] ( Figure 6 ). No significant main effect of Retention schedule was found for any of the frequency ratios [ F (1,38) = 0.001, p = 0.981; F (1,38) = 0.009, p = 0.924 and F (1,38) = 0.313, p = 0.579 for 1∶1, 1∶2 and 2∶3 frequency ratio, respectively]. In addition, there was no interaction of Group × Retention schedule on any of the frequency ratios [ F (1,38) = 1.159, p = 0.288; F (1,38) = 1.308; p = 0.260 and F (1,38) = 0.636; p = 0.430 for 1∶1, 2∶3 and 1∶2 frequency ratio, respectively], indicating again that randomized versus blocked retention testing did not affect performance.
Effect of group on performance persistence
Planned comparisons revealed a significant partial interaction effect from EoA to IR in all 3 frequency ratios [ F (1,38) = 18.929, p <0.001; F (1,38) = 11.064, p = 0.002 and F (1,38) = 5.419, p = 0.025 for 1∶1, 2∶3 and 1∶2 frequency ratio respectively], indicating a differential change in performance between both practice groups from the end of training to immediate retention. Specifically, the randomized practice group showed more skill persistence from EoA to IR in contrast to the blocked practice group ( Figures 4 , 5 and 6 ). Partial interaction effects were also found from EoA to DR in all 3 frequency ratios [ F (1,38) = 7.241, p = 0.011; F (1,38) = 4.264, p = 0.038 and F (1,38) = 4.241, p = 0.046 for 1∶1, 2∶3 and 1∶2 frequency ratio respectively], indicating better performance persistence, from the EoA to DR for the randomized in contrast to the blocked practice group. Post hoc analyses revealed significant performance deterioration from EoA to IR in the blocked practice group for the 1∶1 ( p <0.001) and 2∶3 ( p = 0.004) frequency ratios while performance remained stable in the 1∶2 frequency ratio ( p = 0.984). For the random practice group, performance from EoA to IR remained stable ( p = 1) for all frequency ratios. With respect to long-term retention, the blocked practice group showed significant performance deterioration from EoA to DR in all frequency ratios ( p <0.001 for the 1∶1 and 2∶3 frequency ratios and p = 0.002 for the 1∶2 frequency ratio) while performance of the random practice group remained stable over this one week period ( p = 0.889 for the 1∶1 frequency ratio and p = 1 for the 2∶3 and 1∶2 frequency ratio).
The purpose of this study was to explore the CI effect in a complex bimanual coordination task. First, we hypothesized that a random practice schedule would have detrimental effects during the acquisition phase, but would result in better immediate and delayed retention performance. There was an overall pattern in which the random practice group performed considerably worse at the beginning of the acquisition phase, but with practice, performance of the random practice group progressed towards the performance level of the blocked group. As expected, immediate and delayed retention were superior after random compared to blocked practice, although this effect was absent in the more difficult 1∶2 ratio. Finally, we hypothesized that randomized practice would show more performance persistence during retention intervals, i.e. 5 minutes and one week after the end of practice. This expectation was confirmed in all three frequency ratios.
As stated in the introduction , both the elaboration and action-plan reconstruction hypotheses suggest that enhanced cognitive effort and processing during high CI is responsible for beneficial learning effects. However, none of these hypotheses entail different predictions regarding CI in relation with complex skill learning, in which high attention and memory demands are inherent to the nature of the task itself, and thus requiring more cognitive effort in contrast to simple tasks. We will discuss the effects of CI during complex bimanual coordination task learning with a focus on the temporal evolution of memory processes.
One key feature of the CI effect is that a blocked practice schedule facilitates performance during the acquisition phase compared to a random practice schedule [4] , [33] , [34] . Although our results did not show a main effect between groups, which may be due to the longer training schedule, there was a robust interaction in which performance of the randomized group progressed towards the blocked group in each of the frequency ratio conditions. In line with other studies, better performance of the blocked group was more pronounced early in practice [2] , [22] . As confirmed by previous work, longer practice schedules can even result in the random practice group to outperform the blocked group during training [17] . This suggests that the detrimental effects of random practice during the acquisition phase can be overcome. Due to the presumed higher difficulty level of the 1∶2 frequency ratio, the CI effect could not be overcome during practice. This may imply that it takes more time for the random practice group to stabilize and reach a similar performance level as the blocked group when the difficulty level increases. Some additional points regarding the performance changes in the blocked practice group across the acquisition phase are worth noting. First, performance differences between the first acquisition blocks and initial baseline performance in the blocked practice group are more prominent than appears at first sight. Moreover, performance improvements from baseline to TR16 in the blocked practice group were present. Furthermore, each data point in the acquisition phase (e.g. TR1) already consisted of 6 trials (3 CW and 3 CCW). In addition, only trials without concurrent FB are shown in the figures. Because of the fading feedback schedule, more concurrent FB trials were offered at the beginning of acquisition. As stated above, better performance of the blocked group was clearly more pronounced early in practice [2] , [22] . Maslovat et al. [17] , for example, tested the CI effect while learning a bimanual coordination task by using an extensive practice schedule (100 trials per coordination pattern). Regarding the acquisition data, the authors showed that the blocked practice group did not improve their performance significantly on the 90° coordination pattern and, little performance improvements were present after the first 18 acquisition trials on the 45° coordination pattern. In line with these interpretations, ceiling effects were indeed reached very early in the blocked practice group.
Retention performance was not dependent on the context in which retention was obtained (i.e. blocked or random retention schedule). In this respect, the specificity of learning hypothesis [35] , which predicts that conditions during practice which most closely match the criterion conditions will be most effective for learning that criterion, is not supported here. This is consistent with the study of Shea et al. [22] in which the context of retention had no influence on performance following a random practice schedule. However, in contrast to Shea et al. [22] , retention schedule performance was also not influenced following a blocked practice schedule. Reviews have reported mixed results regarding the magnitude of performance benefit following a random practice schedule [4] , [33] . In the current study, performance differences during retention favoring the random group were confirmed in the 1∶1 and 2∶3 frequency ratios, while no performance differences were observed in retention tests of the 1∶2 frequency ratio. Brady [33] noted that the effect of CI could be a function of the difficulty of a task. In line with this notion, advantages of high CI were found when learning a drawing task; however, the advantage tended to be more pronounced in the simplest versions of the task in contrast to the most difficult version, even though this effect was not significant [20] . Wulf and Shea [5] suggested that there might be a link between the amount of practice and task difficulty. They stated that random practice is more effective when practitioners become more experienced in a complex task, such that the cognitive demands, needed to complete the tasks, are reduced. In line with this notion, random practice in complex tasks may lead to a system overload early in practice, when attention, memory and motor demands are high [5] . The higher memory or motor demands required to perform the 1∶2 ratio, in combination with high CI during practice, may have overloaded the system and, in turn, disrupted the beneficial effects of high CI in this ratio. The CI effect in the 1∶2 frequency ratio may thus be increased by increasing the amount of practice.
Performance persistence
We hypothesized that the random as compared to the blocked practice group would show more performance persistence during the transition from the acquisition to the retention phase. This was confirmed in all three frequency ratios. Moreover, while performance of the random practice group remained stable over a one week period (EoA to DR), performance of the blocked practice group showed a significant deterioration during this time interval. Kantak and Winstein [10] mentioned the importance of the time interval between the end of acquisition and retention tests in order to reveal performance changes, which provides insight into distinct memory processes. A key question here is whether better performance persistence in the random group was a result of more efficient post-acquisition consolidation processes. At first sight, the answer seems to be positive as consolidation is defined as a set of post-acquisition changes wherein a new skill is strengthened [36] . However, these results should be interpreted with caution, as the three distinct memory processes (encoding, consolidation and retrieval) are interrelated and may partially overlap in the temporal domain [10] . Especially in this study, where practice is divided over multiple days, these processes are even more intertwined.
First, it is known that motor performance measured during practice can be related to two main effects of practice: relatively permanent effects, which are conceptualized as learning effects, and temporary or transient effects, which often vanish when the manipulation is removed [37] . Both the elaboration and the action-plan reconstruction hypotheses assume that the greater investment in task-related cognitive processing during random practice will result in the development of a stronger memory representation, which leads to more permanent effects [34] . By contrast, blocked practice will provoke more temporary or transient effects, which are beneficial for acquisition performance. Thus, the faster decrease in performance following the acquisition phase in the blocked group might be explained by (a) a more fragile memory representation as a result of different encoding processes than the random group, (b) the fading of the beneficial temporary effects that are present during practice, or a combination of both explanations. Second, as practice was distributed over three practice days (within a one week period), subjects had the potential to consolidate, and thus strengthen the memory representation in-between practice sessions. As the blocked group practiced only one frequency ratio a day, it was not possible to assess between-practice consolidation processes, which may be a limitation of the present study.
As already stated in the introduction ; studies examining the CI effect in complex motor skills have led to contrary results [5] . Tsutsui et al. [21] examined the effect of CI in learning new patterns of bimanual coordination using multiple days of training. The authors did not find any effect of CI when all coordination patterns were practiced within each practice day. However, when the blocked practice group learned each pattern on separate days, typical CI effects were reported. Albaret and Thon [20] examined the effects of task complexity on CI using a unimanual drawing task. The authors demonstrated retention benefits following random practice; however, the advantage tended to be more pronounced in the simplest versions of the task in contrast to the most difficult. By contrast, the authors did not find any influence of CI when accuracy of orientation, i.e. directional error, was taken into account. Nevertheless, in both papers [20] , [21] , performance stability from end of acquisition to delayed retention was not statistically compared between groups.
Finally, it is important to consider a possible confound in our design regarding different retention delays in the blocked practice group as a result of a fixed practice order. Please note that our experiment was designed this way because we hoped for the best possible learning effects by providing incremental task difficulty. One might argue that the decrement in performance from EoA to IR in the blocked practice group might be mediated by different retention intervals and thus reflects differential forgetting, i.e. more skill deterioration in the 1∶1 frequency ratio (minimum delay of 2 days from EoA to IR) compared with the 1∶2 frequency ratio (5 minute delay from EoA to IR). Therefore, a control experiment (n = 25) was conducted in which frequency ratios were presented in a blocked manner, but counterbalanced over practice days. Subjects were randomly assigned to one of 6 different practice orders (for more details, see File S1 ). Results indicated that the behavioral pattern of the counterbalanced blocked practice group was similar to the fixed blocked practice group described in the current paper. This provides compelling evidence that the effects in the current experiment are not due to different retention intervals.
In summary, we can conclude that random practice resulted in better skill persistence in a complex bimanual coordination task. Specifically, while the blocked practice group showed significant skill deterioration over a one week period, performance of the random practice group remained stable. This effect was evident in all three coordination patterns of various difficulty levels. Although better skill persistence following a random practice schedule was found, the random group could not outperform the blocked group in the most difficult frequency ratio.
The finding that high as compared to low CI led to better skill persistence in a complex bimanual coordination task, even one week after the practice period, is important for future research. Following Kantak and Winstein [10] , we agree that examining the temporal evolution of performance will provide more insight into the mechanisms that implement the learning-performance distinction. In the past, research regarding the CI effect in complex tasks yielded mixed results [5] . However, if we want to provide adequate recommendations for practical settings, we have to examine to what extent the CI effect is generalizable to complex skill learning.
Supporting Information
Overall behavioral pattern. Effect of practice order (fixed versus counterbalanced blocked practice) on error score (ATDlog, i.e. the log-transformed average target deviation) for baseline, acquisition phase (TR1-16), immediate retention (IR) and delayed retention (DR) (mean ± standard error). Better performance is indicated with lower levels of ATDlog.
https://doi.org/10.1371/journal.pone.0100906.s001
1:1 frequency ratio. Effect of practice order (fixed versus counterbalanced blocked practice) on error score (ATDlog, i.e. the log-transformed average target deviation) for baseline, acquisition phase (TR1-16), immediate retention (IR) and delayed retention (DR) (mean ± standard error). Better performance is indicated with lower levels of ATDlog.
https://doi.org/10.1371/journal.pone.0100906.s002
2:3 frequency ratio. Effect of practice order (fixed versus counterbalanced blocked practice) on error score (ATDlog, i.e. the log-transformed average target deviation) for baseline, acquisition phase (TR1-16), immediate retention (IR) and delayed retention (DR) (mean ± standard error). Better performance is indicated with lower levels of ATDlog.
https://doi.org/10.1371/journal.pone.0100906.s003
1:2 frequency ratio. Effect of practice order (fixed versus counterbalanced blocked practice) on error score (ATDlog, i.e. the log-transformed average target deviation) for baseline, acquisition phase (TR1-16), immediate retention (IR) and delayed retention (DR) (mean ± standard error). Better performance is indicated with lower levels of ATDlog.
https://doi.org/10.1371/journal.pone.0100906.s004
Fixed versus counterbalanced blocked practice. In order to test whether the different retention delays in the blocked practice group influenced our results, a control experiment was conducted.
https://doi.org/10.1371/journal.pone.0100906.s005
Acknowledgments
We would like to thank René Clerckx for programming the task.
Author Contributions
Conceived and designed the experiments: LP SPS IAMB. Performed the experiments: LP. Analyzed the data: LP IAMB. Contributed reagents/materials/analysis tools: LP SPS IAMB. Wrote the paper: LP SPS IAMB.
- 1. Battig WF (1979) The flexibility of human memory. In: Cermak LS, Craik FIM, editors. Levels of processing in human memory. Hillsdale, NJ: Erlbaum. 23–44.
- View Article
- Google Scholar
- 3. Magill RA (2011) Motor learning and control: Concepts and applications. New York: McGraw-Hill.
- 6. Shea JB, Zimny ST (1983) Context effects in memory and learning movement information. In: Magill RA, editor. Memory and control of action. Amsterdam: North-Holland 345–366.
- 8. Lee TD, Magill RA (1985) Can forgetting facilitate skill acquisition? In: Goodman D, Wilberg RB, Franks IM, editors. Differing perspectives in motor learning, memory, and control. Amsterdam: Elsevier. 3–22.
- 27. Marteniuk RG (1976) Information processing in motor skills. New York: Holt, Rinehart, and Winston.
- 28. Kay H (1970) Analyzing motor performance. In: Connolly K, editor. Mechanisms of motor skill development. New York: Academic Press.
- 29. Schmidt RA, Lee TD (1999) Motor control and learning: A behavioral emphasis: Human Kinetics.
- 34. Lee TD, Simon D (2004) Contextual interference. In: Wiliams AM, Hodges NJ, editors. Skill acquisition in sport: research, theory and practice. London: Routledge. 29–44.
- Work with Me
- Journal Club
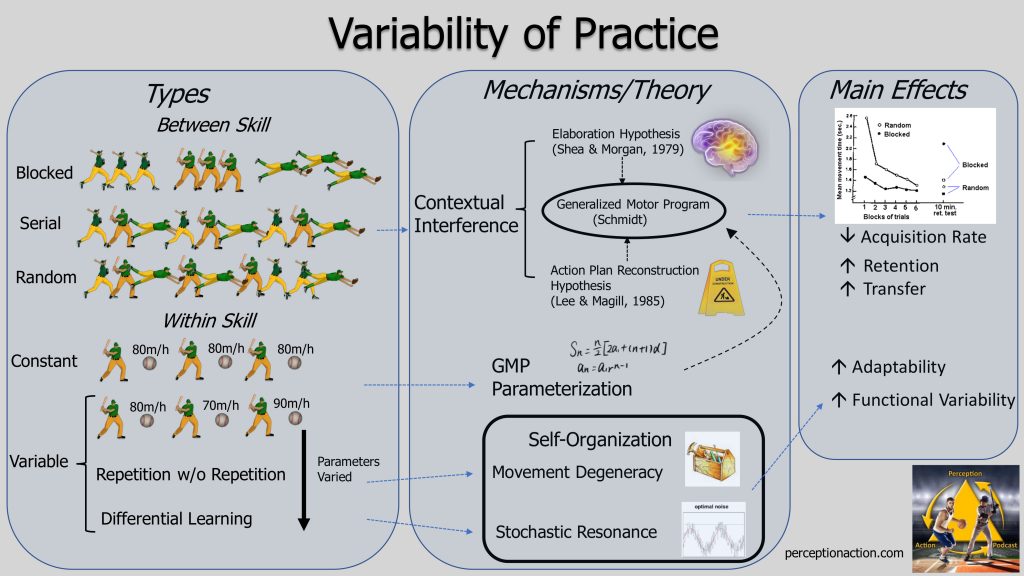
Overview: “Schmidt and colleagues directed attention towards the suggestion that performance on a novel task will be better following varied, rather than constant, prior practice. This suggestion was termed the ‘ variability of practice hypothesis ’ by Moxley (1979) – a label that has since that time become generally adopted. This hypothesis proposes that not all practice is equally efficient in improving motor control; the claim being that variable (i.e. continually changing) practice in contrast to constant (i.e. always the same) practice leads to superior learning”. van Rossum 1990
Key Terminology & Definitions :
– Between-Skill Variability – Switching between different skills within a practice session E.g., Serving, forehands and backhands in tennis OR chipping, putting and driving in golf. There are 3 basic ways between-skill variability can be manipulated in practice:

1) Blocked practice – practicing by performing the different skills in blocks, that is several repetitions of one skill before shifting to the next. E.g., 15 serves followed by 15 forehands followed by 15 backhands.
2) Random practice – practicing by switching between the different skills in a random order such that the order is unpredictable to the athlete. E.g., 1 serve, 2 forehands, 1 backhand, 2 serves, etc.
3) Serial practice – practicing by switching between the different skills in a structured, predictable order. E.g., 1 serve, 1 forehand, 1 backhand, 1 serve, 1 forehand, 1 backhand, etc….
– Within-Skill Variability – Switching between variants of the same skill within a practice session E.g. shooting a jump shot from 10, 15 and 20 feet in basketball OR hitting an iron shot from a flat, downhill and uphill lie in golf. There are 4 basic ways that within-skill variability can be manipulated in practice:

1) Constant practice – practicing by repeating the same variant of the skill over and over eg., 20 iron shots from the same distance and lie.
2) Variable practice – practicing by varying one parameter of the skill from execution to execution (in either a structured or random manner). E.g. switching between different lies in golf, distances in basketball shooting or pitch speeds in baseball batting.
3)” Repetition without repetition ” – practicing be varying multiple parameters of a skill from execution to execution by presenting the athlete with different movement problems to solve E.g., downhill lie from 20 feet, uphill lie into the wind in the rough, etc…
4) Differential learning – practicing by varying multiple parameters of a skill from execution to execution by instructing the athlete to execute the skill in different pre-defined ways E.g. different stance widths, different club lengths, with one eye closed, hands different widths apart on the club etc. **Note: this only one type of DL – see the Differential Learning resource page for more detail.
*Based on these definitions, strict repetition or isolated drilling in practice is blocked, constant practice.
Key Research Findings :
**NOTE #1: The effects we are trying to explain here are “variability of practice” effects NOT “Contextual Interference” effects . As will be discussed below, Contextual Interference is a theoretical explanation proposed to explain variability of practice findings. These two are conflated in many articles.
**NOTE #2: There are also many studies out there that don’t distinguish within and between skill variation so they say “blocked vs random” when they really manipulated “constant vs variable”
1)Blocked vs Random Practice
In their 1979 study, Shea & Morgan asked participants to perform a task of “knocking down barriers” on a table using their hand. Three separate tasks were created by defining 3 different orders in which they had to be knocked down (e.g., eight rear, left middle, and right front). The blocked group performed 18 trials with the same knock down order before switching to a different one. The random group practiced a randomly chosen sequence of knockdown orders (indicated by the “stimulus lights”). After training, both groups also performed 2 transfer tests with new knockdown orders. The dependent measure was the time taken to complete the task. The figure below shows their apparatus and now famous pattern of results:
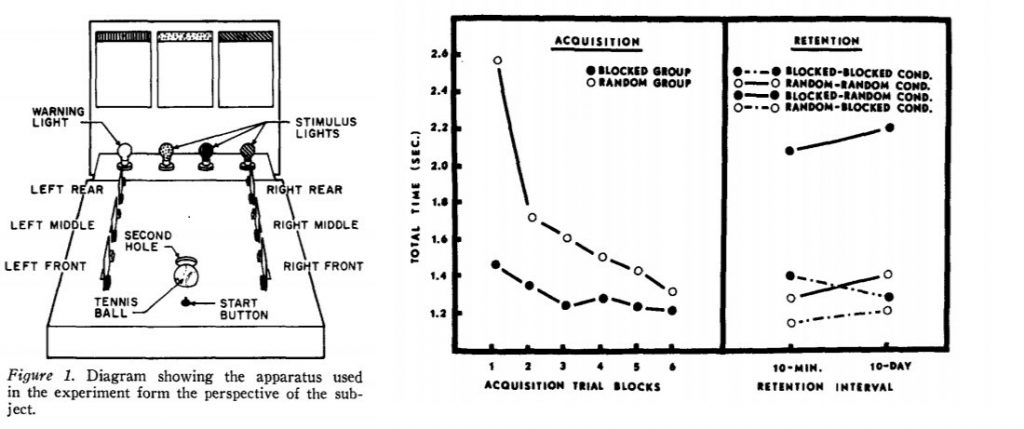
Main Results:
1) Random practice leads to a slower rate of acquisition of the skills as compared to block practice 2) In retention tests, performance for the Blocked group was impaired when asked to now perform the same skills in random conditions 3) Random practice leads to better performance on transfer tests of new skills as compared to Blocked practice (results not shown)
2)Constant vs Variable Practice
Although there have been fewer studies, in general the pattern of results is highly similar: Variable practice leads to slower acquisition but superior retention and transfer to a new task as compared to constant practice. For example, these effects were demonstrated in a study by Shea & Kohl (1990) which required participants to produce movements of different forces and a study by McCraken & Stelmach (1977) which involved variable movement timings.
3)Repetition without Repetition
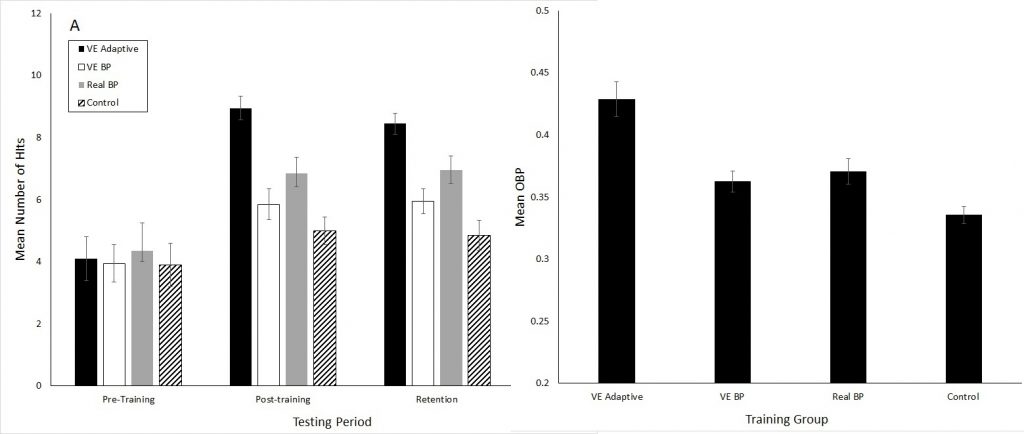
Research comparing constant practice to training conditions in which multiple parameters of a skill are being varied from execution-to-execution has provided evidence that the latter results in greater performance benefits post-training and a higher level of movement degeneracy, as compared to constant practice. Examples include my 2017 baseball batting study which compared an Adaptive Training group (repetition without repetition in the pitch speed, location and type) with two different types of constant practice (always same pitch within a session, either real or virtual training). The repetition without repetition (VE Adaptive) group had both better batting performance in an on-field test and higher OBP in the full season following the training:
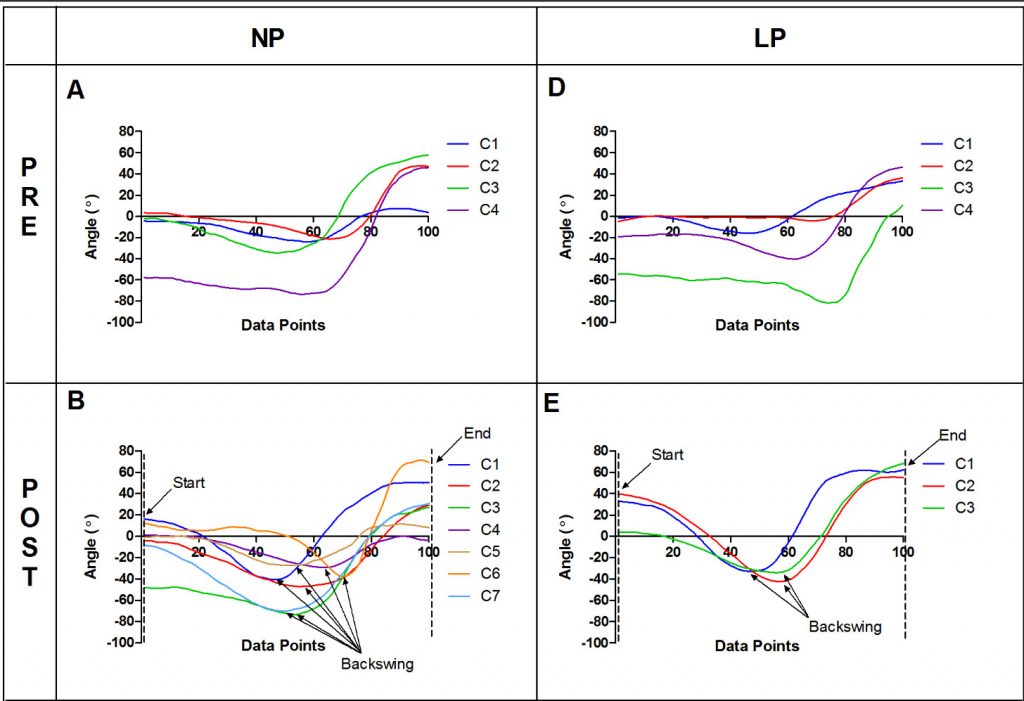
In a 2014 study , Lee et al compared a training group that practiced using repetition without repetition with a group that followed more constant, repetitive practice in tennis. A cluster analysis was used to compare the redundancy in the stroke – more clusters indicated the players were executing the stroke in more different ways. While the number of clusters was similar in the pre-test, there were significantly more clusters for the repetition without repetition group (NP in the figure) in the post-test:
4)Differential Learning
As discussed in detail on this page , a Differential Learning approach of varying multiple parameters has provided evidence for greater post-training performance improvement as compared to traditional, constant practice in sports including speed skating, the shot putt, soccer passing and dribbling, and badminton.
Proposed Theoretical Explanations for Variability of Practice Effects :
The effects of variability of practice can be understood from either an Information Processing or Ecological Approach to skill acquisition .
– The Information Processing Explanation: Contextual Interferen ce & Re-parameterization
The most often discussed theoretical explanations for the differences between blocked vs random practice and constant vs variable practice are based on Schmidt’s Schema Theory of Motor Control . In this theory, our actions are controlled by Generalized Motor Programs (GMP). A GMP contains an abstract representation with invariant features pertaining to the order of events, the relative timing of events and the relative force with which events are produced. In other words it is the basic technique required for a movement stored in an internal model in memory. In order to determine how a particular movement should be performed the specific parameters such as overall movement duration, overall force of contractions and the muscles involved are specified to the GMP.
Within this approach, Random practice is thought to induce Contextual Interference . That is, when a performer is developing a GMP for one skill (a serve in tennis) switching to practicing another skill (a backhand) right away will interfere with the development process. The presence of this interference explains why acquisition of the skill is slower under conditions of random practice compared to blocked . The longer-term benefits of random practice (seen in retention and transfer tests) have been explained in terms of two different possible mechanisms:
1) The Elaboration Hypothesis (Shea & Morgan, 1979)
In the Elaboration Hypothesis , it is proposed that random practice allows a performer to develop more elaborate and distinct motor programs. For example, because in random practice a tennis player would be frequently switching back and forth between forehands and backhands, this would allow them to better compare and contrast the two movements, identify the key differences, and develop more distinct GMPs. More elaborate GMPs are thought to allow for better comprehension/understanding of the skill and quicker retrieval of the GMP from memory. In this hypothesis, the comparison between tasks is thought to occur during the interval between the end of the last execution and the start of the next one.
2)The Action Plan Reconstruction Hypothesis (Lee & Magill, 1985).
In the Action Plan Reconstruction Hypothesis (also sometimes called the Forgetting-Reconstruction Hypothesis ), it is proposed that random practice forces a performer to abandon (or forget) their action plan (GMP) with each new trial, because they are likely moving to executing a different skill. This required forgetting means that the next time they have to perform that same movement again, they will need to reconstruct the plan they used last time. So for example, on trial #1 I execute a forehand with GMP A. Now on trial #2 I have to do a backhand using GMP B, so I need to forget GMP A. Then on trial #5, when I have to do another forehand, I need to re-assemble GMP A again. This continuous forgetting-reconstruction process is thought to lead to a stronger representation of the task in memory allowing for faster retrieval.
Within all of this is the idea that different amounts of variability in practice will lead to different amounts of Contextual Interference. So interference increases as you go from Blocked- Serial -Random Practice, for example.
– Variable Practice & Parameterization
Within this Schema Theory approach, the benefits of variable practice over constant practice can be understood in terms of parameterizing the GMP. For example, switching the distances or lies of a golf shot from swing to swing requires re-parameterization (setting different forces, durations, timings, etc in the GMP). Thus, after training, the performer can do this parameterization process more quickly and accurately following variable practice. I like to call this developing Adjustability, as I described in this post looking at variability .
– The Ecological Approach Explanations: Movement Redundancy & Stochastic Resonance
In the Ecological Approach, instead of developing adjustability through variability, the benefits of variability of practice are proposed to arise through the development of Adaptability (i.e., developing the ability to solve movement problems in different ways). This comes through two different proposed mechanisms:
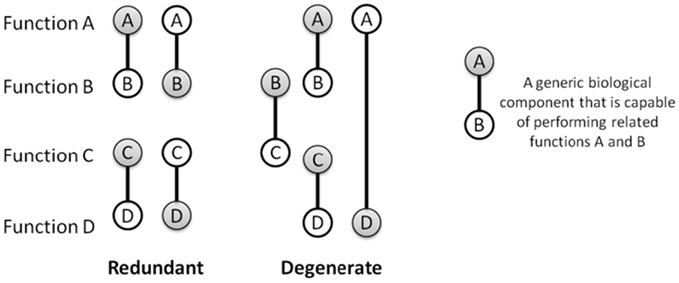
1)Movement Degeneracy
Degeneracy , which occurs in large number of biological systems, is the ability of elements that are structurally different to perform the same function or yield the same output. So, for example, in volleyball I can serve the ball using just my shoulder, my shoulder and elbow or my shoulder elbow and wrist, etc. Degeneracy leads to a more robust and adaptable system because if one way of doing it doesn’t work you can switch to others. Varying a large number of movement parameters (repetition without repetition) is thought to promote the development of degeneracy because it is likely the performer will not be able to use the same movement solution for each new problem you create.
2)Stochastic Resonance
In the theory of Differential Learning it is proposed that adding variability to practice conditions improves performance through a process called stochastic resonance. As explained in detail on my DL resource page , this is the idea that noise (the practice conditions) can be constructive in that it can actually help to strengthen a weak signal (the athlete’s movement pattern). From this viewpoint, blocked or constant practice are likely to have too low amount of noise to promote stochastic resonance.
Some Qualifications to the Classic Findings & Moving Beyond Categories of Variability
As summarized nicely in a chapter published by Farrow & Buszard in 2017 (discussed in podcast episode #91 ), there are some important limitations to applying the classic variability of practice effects in coaching. The first issue concerns complexity. In the bulk of the research on variability of practice, very simple skills with relative few degrees of freedom have been used, for example tapping out sequences with your fingers on a keyboard. Research which has examined more complex motor skills like those involved in most sports have found much smaller differences between random and blocked practice. For example, in the meta-analysis conducted by Brady in 2004 , the average effect size (ES) found in basic, laboratory research studies was .57, while it was only .19 for applied research. For reference, 0.5 is considered to be medium ES while 0.2 is small. It is also the case that cleanly dividing practice into blocked vs random, constant vs variable is not as easy in real world settings. Finally and maybe most importantly, Brady found that age and experience strongly mediate the benefits of variability of practice with random and variable practice being less beneficial for new learners :
These experience effects have led some researchers (e.g. Porter & Beckerman, 2016 ) to propose that transitioning from blocked to random practice or using serial practice (which has some elements of both) may be more effective for novices and children.
Moving Beyond Categories
In light of these limitations, Farrow, Buzzard and colleagues have proposed that it may be more beneficial to use a continuous metric (which captures both within and between skill variations) to quantify and manipulate practice variability rather than having distinct categories. An example of this concept illustrated for tennis is shown below:
Summary & Conclusions:
No matter which way you slice it or which skill acquisition approach you align yourself with, there are clear, unequivocal benefits to adding variability to practice. To me this is the lowest hanging fruit that still remains in a lot of the practice sessions I observe. However, it is also important to note the the amount of variability you add needs to be appropriate for the skill level and age of the athlete you are coaching. Variability added by practice conditions is going to interact with the variability inherent in the performer themselves (which is generally higher for new learners). So, less variability of practice will typically be better.
– Schmidt’s schema theory: The empirical base of the variability of practice hypothesis: A critical analysis – Contextual interference effects on the acquisition, retention, and transfer of a motor skill – Contextual interference: a meta-analytic study – Exploring the applicability of the contextual interference effect in sports practice – Time scales of adaptive behavior and motor learning in the presence of stochastic perturbations – Changing Up the Routine: Intervention-Induced Variability in Motor Learning – Neurobiological degeneracy: A key property for functional adaptations of perception and action to constraints
Podcast Episodes :
– 9 – Off the Learning Curve: Measuring Skill Acquisition – 91 – Variability of Practice Revisited – 100 – The Why, What, How & When of Movement Variability – 121 – How Much Variability in Practice & When? Does Water Immersion Enhance Motor Learning?

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
Interleaved practice benefits implicit sequence learning and transfer
Julia m. schorn.
Department of Psychology, University of California, Los Angeles, Los Angeles, CA 90095 USA
Barbara J. Knowlton
Compared to blocked practice, interleaved practice of different tasks leads to superior long-term retention despite poorer initial acquisition performance. This phenomenon, the contextual interference effect, is well documented in various domains but it is not yet clear if it persists in the absence of explicit knowledge in terms of fine motor sequence learning. Additionally, while there is some evidence that interleaved practice leads to improved transfer of learning to similar actions, transfer of implicit motor sequence learning has not been explored. The present studies used a serial reaction time task where participants practiced three different eight-item sequences that were either interleaved or blocked on Day 1 (training) and Day 2 (testing). In Experiment 1 , the retention of the three training sequences was tested on Day 2 and in Experiment 2 , three novel sequences were performed on Day 2 to measure transfer. We assessed whether subjects were aware of the sequences to determine whether the benefit of interleaved practice extends to implicitly learned sequences. Even for participants who reported no awareness of the sequences, interleaving led to a benefit for both retention and transfer compared to participants who practiced blocked sequences. Those who trained with blocked sequences were left unprepared for interleaved sequences at test, while those who trained with interleaved sequences were unaffected by testing condition, revealing that learning resulting from blocked practice may be less flexible and more vulnerable to testing conditions. These results indicate that the benefit of interleaved practice extends to implicit motor sequence learning and transfer.
Introduction
From the moment we wake up to the moment we fall asleep, we perform many skills that we have learned over time, like brushing teeth, typing, driving a car, or playing a musical instrument. Complex and simple skills alike rely on motor dexterity, sequence learning, perceptual acuity, and both explicit and implicit learning (Shmuelof & Krakauer, 2014 ). Procedural skill learning is a persistent and crucial part of the human experience, so determining an optimal practice schedule is essential as we are heavily dependent on our ability to learn new skills throughout our lifetime. Practice schedules that introduce high contextual interference (CI) by interleaving, or randomizing, tasks hinder initial performance but aid in long-term skill retention. While the CI effect is robust, it has yet to be reliably demonstrated in implicit motor sequence learning and transfer. Determining if the CI effect persists in the absence of explicit knowledge is essential in order to design effective practice schedules, especially for clinical populations who demonstrate impaired explicit memory but preserved implicit memory. For example, patients with amnesia show intact motor sequence learning but impaired declarative memory (Reber & Squire, 1994 ). Optimizing practice schedules to enhance long-term retention and transfer of learning will benefit lives across a broad spectrum of settings.
Decades of cognitive psychology research have demonstrated that CI through interleaved, or random, practice (compared to blocked practice) is a way to effectively acquire skills that can be retained in the long-term (Battig, 1966 ; Magill & Hall, 1990 ; Shea & Morgan, 1979 ). Interleaving tasks or stimuli can be thought of as a “desirable difficulty” as it hinders initial performance but results in superior long-term retention and transfer performance (Bjork, 1994 ). Blocked practice, on the other hand, facilitates acquisition performance because it requires the learner repeatedly performs a single task before moving on to the next one. In one of the earliest demonstrations of the benefits of CI in motor skill learning, subjects practiced three sequential arm movement tasks, which were either blocked together or intermixed in an interleaved condition (Shea & Morgan, 1979 ). While performance was worse in the interleaved condition during practice, subjects showed better retention and transfer to similar tasks. Blocked practice usually facilitates faster acquisition but results in poorer long-term retention. This differential effect exemplifies the difference between learning and performance, a critical distinction (for a review, see Soderstrom & Bjork, 2015 ). For example, a pianist may quickly master a difficult passage by practicing it repeatedly only to forget all progress the next day, because conditions that enhance performance may not enhance learning.
At first glance, the CI effect seems to be at odds with the Specificity of Learning hypothesis, which posits that learning is most optimal when the practice conditions during the acquitision phase are the same as those during the testing phase (Barnett et al., 1973 ). Some findings in the motor learning literature suggest there is partial support for this hypothesis, though the CI effect is more robust (Shea & Kohl, 1990 ; Travlos, 2010 ). Thus, interleaved practice shows benefits even when tasks are blocked at test.
Theorized mechanisms of the contextual interference effect
Though the benefits of contextual interference are well documented, the mechanism by which interleaving facilitates this enhanced retention is still not completely understood. Two general hypotheses currently stand out. The forgetting-reconstructive hypothesis (or action plan reconstruction hypothesis) posits that interleaving is beneficial because it requires each task set to be frequently retrieved, while during blocked practice the task set remains in working memory for the entire task duration (Lee & Magill, 1983 ; Lee et al., 1985 ). When practicing interleaved motor sequences, a pattern must be learned and then immediately “dumped” from working memory in order to prepare for subsequent trials (Lee & Simon, 2004 ). In each trial, the learner must retrieve a motor pattern into working memory or construct one from scratch. Conversely, blocked motor sequences can remain in working memory for multiple trials without needing to be updated. While frequently forgetting and retrieving stimuli may initially hinder performance, it also allows the learner to practice reconstructing motor patterns, which is beneficial for long-term retention. This hypothesis may partially explain the dissociation between learning and performance often seen in the CI effect (for a review, see Kantak & Winstein, 2012 ).
On the other hand, the elaboration-distinctiveness hypothesis (or discriminative-contrast hypothesis) posits that interleaved practice facilitates organizational and item-specific processing, so that subjects frequently compare different stimuli for more durable encoding (Shea et al., 1985 ; Shea & Zimny, 1983 ). When tasks or stimuli are interleaved, differences between them may be more easily discerned as compared to when they are blocked.
Though traditionally pitted against one another, these two theories are not mutually exclusive and may explain different components of the CI phenomenon (Lin et al., 2008 ). In an fMRI study, interleaved practice of a motor sequence task compared to blocked practice resulted in increased activation in sensorimotor regions, followed by a decreased activation in similar regions during a delayed retention test (Lin et al., 2011 ). The increased activation during encoding of interleaved sequences was interpreted as representing additional motor program reloading. Since there was increased retrieval processing during encoding, retrieval at test required less activation in these regions. This view lends credence to the forgetting-reconstructive hypothesis.
However, it appears that interleaving may also offer greater benefits than simply requiring more retrieval. In a study where participants learned to identify paintings by artist in either a blocked or interleaved fashion to develop a concept of different artists’ styles, participants in the interleaved condition showed better learning than those who studied the paintings in a blocked but spaced schedule (Kang & Pashler, 2012 ). As temporal spacing of study items required explicit retrieval of the artist’s names, inferior performance in the blocked condition indicated that retrieval practice alone cannot account for the benefits of interleaving. Participants had to abstract information across paintings to learn artists’ styles that could be used to classify new paintings, so the contrasts and comparisons made at encoding in the interleaved condition may have been important for generalization, supporting the elaboration-distinctiveness view.
Neither of these theories specifically account for implicit processes important for motor skill learning, and research on the benefits of interleaving has largely ignored the interplay between implicit and explicit learning (Bjork & Kroll, 2015 ; Shanks & St. John, 1994 ). In light of this, an alternate theory has emerged that proposes that high CI and increased task switching overloads an individual’s working memory capacity, preventing them from gaining explicit task-relevant knowledge (Rendell et al., 2011 ) and enabling greater implicit learning. Previous research showing increased levels of cognitive activity with interleaved practice as compared to blocked practice lends some support to this theory (Li & Wright, 2000 ). Recent findings extend this idea, suggesting that high cognitive effort seen in interleaved practice may be partially due to increased error processing as well as task-switching (Broadbent et al., 2017 ). Following an error, greater cognitive activity could be attributed to an individual updating and correcting a rule as well as retrieving information for the upcoming task. However, evidence supporting this hypothesis is sparse and relegated to perceptual-cognitive skills. It is also unclear whether CI simply enables more acquisition of implicit knowledge, or if it leads to implicit knowledge that is better retained and better able to support transfer to similar tasks. In the present study, implicit learning under blocked and interleaved conditions will be compared directly.
The contextual interference effect and implicit motor sequence learning
Research concerning the CI effect and implicit motor learning has largely focused on gross motor skills like those used when playing sports (French et al., 1990 ; Goode & Magill, 1986 ; Menayo et al., 2010 ). Furthermore, research investigating the CI effect in fine motor sequence learning has almost exclusively focused on explicit memory (Wright et al., 2016 ). Though previous research has explored the effect of CI in implicit motor learning, few studies specifically investigate fine motor sequence learning over a substantial delay (Dang et al., 2019 ; Sekiya, 2006 ) and is thus a primary aim of this paper. One such study examined the CI effect in a pursuit-tracking task (Sekiya, 2006 ). The experimenters told participants in the explicit group the presentation order and number of patterns in the task, while those in the implicit group received no instruction. Interestingly, they failed to replicate the CI effect and found no differences between implicit and explicit learners. One possibility for this finding is the relatively high CI in the Blocked group may have reduced differences between the practice conditions. Blocked segments were intermixed with random segments so that participants did not become aware of the repetition. Thus, it is not yet clear if implicit and explicit learning are similarly affected by CI during practice.
Implicit motor sequence transfer
Positive transfer to novel tasks or contexts is a crucial goal in many training situations as one often cannot train on every possible task variation or in every possible context. It is suggested that when positive transfer occurs, a memory representation of the skill has been created that is more general than a representation that could only support the practiced sequence. Positive transfer would indicate that participants learned not only the practiced sequences but also generalized knowledge that benefits the performance of new sequences. Learning may also be sequence-specific, with performance of new sequences similar to performance at the beginning of practice on the original sequences. However, learning could also result in negative transfer with performance of new sequences impaired due to interference (Obayashi, 2004 ). In this case, learning may be sequence-specific, and this sequence knowledge may impair the ability to perform similar sequences.
Though the benefits of interleaving on retention are well-studied, there is currently less evidence that interleaving can also lead to improved transfer to similar actions (Bangert et al., 2014 ; Brady, 2004 ; Meira & Tani, 2001 ; Russell & Newell, 2007 ; Schmidt & Lee, 2005 ). Transfer in the skill learning domain has been extensively studied, including transfer of learning from one effector to another (Kelso & Zanone, 2002 ), such as right hand to left hand, or scaling, such as performing a skilled action at a different rate or using greater force (Newell, 1996 ). However, there are limited transfer studies on fine motor skills, and none that specifically consider implicit sequence learning over a long delay. Müssgens and Ullén ( 2015 ) showed that interleaved sequences, as compared to blocked, led to better transfer to a new sequence on an immediate test, but it is unlikely that these sequences were implicitly learned as a majority of subjects reported some sequence awareness. Additionally, the immediate test made it likely that interference occurred on the test with the sequences that had just been practiced. Similarly, interleaving may reduce negative transfer of motor sequences when new sequences are performed (Shimizu et al., 2016 ). However, transfer was tested at the end of the experiment, and not after a one day delay when the effects of contextual interference are most apparent. This delay is also crucial for observing “offline gains” in which motor memory is stabilized and improved in the absence of practice and is influenced by sleep (Nader et al., 2000 ; Walker et al., 2002 ). It was also unclear whether sequence knowledge was primarily explicit or implicit in Shimizu et al. ( 2016 ).
Other studies concerning the CI effect in novel motor skill learning found that prior experience with interleaved practice improved new motor task acquisition, however, delayed retention was unaffected by practice schedule (Hodges et al., 2014 ; Kim et al., 2016 , 2018 ). Hodges et al. ( 2014 ) found that random practice experience, compared with blocked practice, led to better acquisition of three novel motor skills after a one-day delay and mitigated the low accuracy cost usually associated with random practice, but ultimately did not affect retention. Using a similar paradigm, Kim et al. ( 2016 ) found that interleaved practice of a motor skill was beneficial for novel skill acquisition, but afforded no retention benefits after a delay. However, interleaved practice in this experiment was not truly random and was more analogous to a blocked practice schedule, as participants practiced 5–15 repetitions of the same motor sequence in a 30-s trial. Kim et al. ( 2018 ) sought to address this limitation by using the discrete sequence production (DSP) task to induce high levels of contextual interference and found that acquisition of a novel task was better for individuals with prior interleaved rather than blocked practice, and this benefit remained after a significant delay. Notably, awareness for the sequences was not probed in any of these experiments and sequence learning was presumed to be explicit. Though participants may not always possess explicit, in-depth knowledge of the task, the DSP task is considered to be an explicit sequence learning paradigm and participants are often informed that they are performing fixed motor sequences (Abrahamse et al., 2013 ; Bo & Seidler, 2009 ). In light of this, our second experiment aims to examine if contextual interference can aid in positive transfer of novel motor sequences when learning is implicit. Demonstrating the CI effect in transfer of sequence learning to different (but similar) sequences would show that this effect persists despite a high degree of potential interference, similar to real-life learning.
The current experiment
The proposed study aimed to add to the body of research on contextual interference by examining the effect of interleaved practice on implicit learning of sequences in a serial reaction time task (SRTT). In this task, participants can acquire knowledge of a sequence of locations as shown by more rapid responses to locations presented in sequence compared to those presented in a random order (Nissen & Bullemer, 1987 ). Participants can show sequence-specific improvement without conscious awareness that the locations appeared in any sequence. Additionally, participants might show non-sequence-specific learning, in that learning and performance of new sequences may be faster than initial performance. Though the SRTT typically examines learning that is specific to the practiced sequence, we argue that learning that generalizes to new sequences may be more relevant to real-world skill learning. In both experiments, training and testing comprised of blocked and interleaved practice conditions, allowing us to examine transfer-appropriate processing (TAP) effects (Morris et al., 1977 ). TAP effects are related to the Specificity of Learning hypothesis which states that performance is optimal when the context during testing resembles the context during training (Barnett et al., 1973 ). There is some support of this hypothesis in the motor learning literature; however, the CI effect is more robust and does not always align with this principle, in that test performance in either condition is often better when preceded by interleaved practice (Shea & Kohl, 1990 ; Travlos, 2010 ).
General method
We examined the effect of practice schedule on motor sequence learning and transfer by either blocking or interleaving the presentation of three different eight-item motor sequences in a two-day experiment utilizing the SRTT. Both experiments thus had four groups, based on condition, for Day 1 (Train) and Day 2 (Test): Blocked training/Blocked test (BB), Interleaved training /Interleaved test (II), Blocked training/Interleaved test (BI), and Interleaved training/Blocked test (IB). At the end of the experiment, explicit knowledge of the sequences was probed with a questionnaire.
Behavioral task
The SRTT has frequently been used to measure implicit learning (Keele et al., 2003 ; Robertson, 2007 ). In this simple task, the participants are asked to respond to cued locations using keypresses. The participant is instructed to respond to the successive locations as quickly and accurately as possible and is not informed that there is a structure governing the order of the appearance of the cued locations. Despite the lack of awareness of the structure, reaction time (RT) is faster for sequences that were practiced compared with RT for a random presentation of cued locations. RT is the primary dependent measure because error rates are generally low and accuracy is not a useful measure of motor learning in this task. This task may share many features of real-world skills that engage fine motor circuits in which movement components must be produced in a specific sequence, such as typing or playing a musical instrument.
Study design
We utilized a between-subjects design with the SRTT. Subjects sat with four fingers of the right hand on four keys on a keyboard (U, I, O, P) that corresponded to the four outlined, unfilled circles on a blue computer screen in a darkened room. One of the circles turned white to act as a cue to press the corresponding key (i.e., the first circle on the screen corresponds with “U”). After the button press, another circle turned white and the first circle reverted to being unfilled. A tone sounded if a subject failed to press a button or if a subject made an incorrect button press. RT and accuracy were recorded for each key press. No baseline performance was measured, since this task is simple and baseline performance does not reliably capture individual differences in learning (Stark-Inbar et al., 2017 ).
Participants practiced three different eight-item sequences that were either interleaved or blocked on Day 1 (training) and Day 2 (testing) (Fig. (Fig.1). 1 ). To control for specific item effects, the sequences were randomized so that no two participants had the same ones. Each sequence contained each letter (U, I, O, P) twice. Critically, participants were not told that there were sequences, only to respond to each cue as quickly and as accurately as possible. Participants were randomized into the Interleaved or Blocked training groups and were tested in either a Blocked or an Interleaved condition, counterbalanced across training conditions. Participants were not told which condition they were assigned to on either day. In the blocked condition, participants received 80 repeated presentations of each sequence (i.e., AAA…BBB...CCC). In the interleaved condition, they received three sequences interleaved for a total of 240 trials (i.e., ACBABCBAC....). This number of trials is comparable to the training procedure used in our previous work (Lin et al., 2010 ).

Sequences and study design. ( A ) Six example sequences. In Experiment 1 , participants learned three sequences; in Experiment 2 , participants learned six sequences – three on Day 1 and three novel sequences on Day 2. ( B ) Study design for both experiments. Day 1 is the same for both while Day 2 is separated out by experiment. Each number represents an eight-item sequence as shown in ( A )
Day 2 was the same as Day 1, participants were randomly assigned to either the blocked or the interleaved condition. In Experiment 1 , the sequences presented on Day 2 to each participant were the same as those presented on Day 1. In Experiment 2 , the sequences presented on Day 2 were novel, so participants practiced three sequences on Day 1 and three different sequences on Day 2, for a total of 6 unique sequences. Each day in each experiment contained the same number of trials.
To assess whether subjects had acquired explicit knowledge of the sequences, a free-recall questionnaire based on past research was administered after the second session (Robertson et al., 2004 ; Willingham & Goedert-Eschmann, 1999 ). Though free recall may also reflect a degree of implicit memory, that is also true of most explicit awareness tests (Shanks & St. John, 1994 ). Though there is no consensus on the best way to measure awareness in this task (Robertson, 2007 ), free recall, rather than recognition, was assessed as it can be argued that it is the most appropriate measure of explicit memory (Frensch & Rünger, 2003 ). The test was comprised of the three following questions, which prompted the participants to recall the sequences:
- Did you notice any pattern(s)?
- How many sequences where there?
- Please type in all sequences (using the same keys) and hit Enter after each sequence.
Participants were also asked how many hours of sleep they got in between Day 1 and Day 2. Sleep has been shown to be critical for offline gains in implicit learning, however, rapid consolidation of a motor skill can occur within hours or even seconds after practice (Bönstrup et al., 2019 ; Kim & Wright, 2020 ; Squire et al., 2015 ; Walker et al., 2002 ). We chose a 24-h interval between training and testing to measure long-term retention as this is sufficient to observe consolidation; subsequent nights of sleep (e.g., 48–72 h from learning) seem to maintain the memory and prevent it from decay, with only minimal performance gains, if any at all (Rickard et al., 2008 ; Walker et al., 2003 ). There is no consensus on the optimal delay between training and delayed retention testing, but this length of delay is typically used as it is a more practical way to assess consolidation (Kantak & Winstein, 2012 ). Contextual interference effects are also more apparent after this length of delay (Cahill et al., 2001 ; Perez et al., 2005 ).
Monte Carlo simulation
The implicit-explicit memory distinction may lie on a continuum, with participants having varying amounts of explicit knowledge. However, since we were primarily interested in purely implicit learners, we dichotomized our sample and post hoc sorted participants based on a cutoff determined by a Monte Carlo simulation a priori. This allowed us to determine the number of sequential elements that would be recalled by chance. Subjects would be considered fully implicit learners if their recall was near chance level, and subjects would be considered as having some explicit sequence knowledge if they exceeded this level. This procedure is similar to past research with the SRTT in which a cut-off score based on chance performance was applied to participants’ item recall to define implicit and explicit groups (Robertson et al., 2004 ; Willingham & Goedert-Eschmann, 1999 ). Like the present study, this was to ensure that the individuals in the implicit group had little to no awareness of the sequence.
To determine the amount of explicit knowledge for practiced sequences, we created a score for each participant based on their percentage correct recall for each sequence and averaged the three scores, with 1 as a perfect score. For example, if a subject remembered four sequential items of one sequence (0.5), two sequential items of the second sequence (0.25), and none of the third sequence (0), those three scores would be averaged to create a subject’s “explicit score.” In this example, this subject would have a score of 0.25, meaning that on average, they remembered two items per sequence.
Participants were considered to have only implicit knowledge of practiced sequences if their explicit knowledge of the sequences was near chance levels. To determine chance levels in terms of the number of sequential elements produced, we ran a Monte Carlo simulation. We compared three randomly generated “test” sequences to a set 1,000,000 randomly generated sequences. The test sequences and the comparison set of sequences were in the same format as used in the experiment (i.e., eight item sequences, using each character (U, I, O, P) twice). The percentage similarity score for each test sequence was calculated averaging across all items in the comparison set. We ran this simulation 10,000 times. The generated percentage scores all fell within 0.23–0.26. Thus, on average, we concluded that chance level of performance would be producing about two sequential items per sequence (.25). We considered a participant to have minimal knowledge of the sequences if the participant was able to produce three or fewer sequential items per sequence and used this a-priori cut off to dichotomize our sample. Implicit learners were participants who recalled, on average, 0–3 sequential items per sequence (at chance) while explicit learners recalled 4 or more items per sequence (above chance).
Experiment 1
Participants.
100 right-handed young adults were enrolled in the study (88 female; age 18–48 years, M = 20.9, SD = 4.2). Participants were undergraduate students recruited from UCLA and were given course credit for their participation. All participants gave informed consent using an institutionally approved consent form. Participants were excluded if they had a history of neurological or psychiatric disease or if they were taking neuroactive medication that could affect sensory processing, movement, or cognition. Since the SRTT is a simple task, accuracy likely reflects the degree of participants’ effort, not learning. Thus, participants were excluded if they had an accuracy lower than 80% on either day, in line with previous research (Willingham et al., 2000 ). All participants reported they they were right-handed, although the degree of handedness was not assessed. A total of 17 participants were excluded for either low accuracy ( n = 8), computer error ( n = 5), or failing to complete the experiment ( n = 4). Our final subject pool consisted of 83 right-handed young adults (59 female, age 18–43 years, M = 20.6, SD = 3.2).
Data analysis
Sequence RTs were calculated by summing the eight key press RTs for correct sequences. In line with previous research, only accurate sequences were analyzed (Reber & Squire, 1998 ). We took an average of the last ten sequence RTs per sequence (A, B, and C) for the blocked training condition, for a total of 30 trials. For the interleaved training condition, we took an average of sequence RTs from the last 30 trials. For the blocked testing condition, we used the same procedure but looked at the first ten trials for each sequence, for a total of 30 trials. Similarly, for the interleaved testing condition, we studied the first 30 trials. To measure retention, difference scores were calculated by subtracting the RT of the last 30 trials from Day 1 from the RT of the first 30 trials from Day 2. A negative difference score, reflecting a decrease in RT, indicates improvement (faster performance) from Day 1 to Day 2, while a positive difference score, reflecting an increase in RT, indicates a decline in performance (slower performance) from Day 1 to Day 2. A difference score of zero would indicate successful retention. We assessed learning during the practice phase by looking at sequence RTs over Day 1 (training) in both implicit and explicit learners using the Mann-Kendall test, a nonparametric test for monotonic trends. We also calculated a learning difference score to assess performance at the beginning and end of training. Here, we compared the first 30 trials of interleaved sequences to the last 30 trials, and the first ten of each blocked sequence the last ten of each. Difference scores were calculated similarly to past research (Lin et al., 2010 ; Wymbs & Grafton, 2009 ). We conducted one-sample t-tests for all groups (BI, II, BB, IB ) to assess if difference scores were significantly different from zero, which would indicate an improvement or a decline in performance from Day 1 to Day 2.
To examine whether the different practice conditions yielded different levels of sequence awareness, we compared the average number of sequence items that participants recalled using an independent t-test. We conducted a 2 × 2 × 2 between-subjects ANOVA, after post hoc sorting subjects into implicit and explicit learners based on their recall score. Training condition (Interleaved, Blocked), testing condition (Interleaved, Blocked), and learner type (Implicit/Explicit) were the three factors while the difference score was the dependent measure. In addition, we conducted an ANCOVA with recall score as the covariate before sorting participants into implicit or explicit learners. Like the ANOVA, training and testing condition were the two factors and the difference score was the dependent measure.
On Day 1, a Mann-Whitney test indicated that participants who practiced interleaved sequences were significantly less accurate ( M = 92.34, SD = 4.38) than participants who practiced blocked sequences ( M = 94.22, SD = 4.13; U = 1134.50, p = .013). However, on Day 2, there was no significant difference in accuracy between those who performed interleaved sequences ( M = 94.02, SD = 4.27) and those who performed blocked sequences ( M = 95.59, SD = 2.51; U = 992.50, p = .229). Those who were tested on interleaved sequences either received blocked or interleaved training the day before, however training condition did not impact accuracy on Day 2 ( M II = 93.34, SD II = 4.44; M BI = 94.74, SD BI = 4.06; U = 280.5, p = .234). Similarly, training condition did not impact accuracy on Day 2 for those who were tested on blocked sequences ( M BB = 95.67, SD BB = 2.32; M IB = 95.52, SD IB = 2.72; U = 197.50, p = .968).
Participants who received blocked practice explicitly recalled on average more sequential items per sequence ( M = 4.18, SD = 2.48) than those who had received interleaved practice ( M = 3.17, SD = 2.13); t (81) = -1.996, p = .049). Though 84.3% of participants reported noticing a pattern, only 30.1% of participants correctly noticed there were three sequences. Before categorizing participants into implicit or explicit learners, we conducted a two-way ANCOVA to control for recall score. We found a significant main effect of training condition ( F (1,78) = 38.06, p < .001), a significant main effect of testing condition ( F (1,78) = 10.895, p = .001), and a significant interaction after controlling for recall score ( F (1,78) = 11.565, p = .001). The covariate was not significantly related to performance, indicating that a participants’ knowledge about the sequence had no significant impact on performance or the benefit of interleaved practice ( F (1,78) = 3.02, p = .08). Since our original interest was purely implicit motor sequence learning, we then separated groups based on a cutoff score denoting chance performance. Learners were sorted into two groups based on their explicit recall score: implicit ( n =40) and explicit ( n =43). See Table Table1 1 for breakdown of individual group n s.
Number of participants per condition and learner type for Experiment 1
| Condition | Implicit Learners | Explicit Learners | Total |
|---|---|---|---|
| II | 14 | 8 | 22 |
| BB | 7 | 12 | 19 |
| IB | 9 | 12 | 21 |
| BI | 10 | 11 | 21 |
| Total | 40 | 43 | 83 |
Notes. Since Implicit/Explicit sorting was determined post hoc, there are uneven numbers in each group. On average, Implicit Learners recalled an average of 1.64 items per sequence ( SD = 1.32) while Explicit Learners recalled an average of 5.54 items per sequence ( SD = 1.28).
II Interleaved/Interleaved, BB Blocked/Blocked, IB Interleaved/Blocked, BI Blocked/Interleaved
For explicit learners, we found significant decreasing monotonic trends for both the interleaved training group ( τ = -.242, p < .0001) as well as the blocked training group ( τ = -.442, p < .0001) (Fig. (Fig.2). 2 ). Thus, both groups showed improvement on the SRTT during training on Day 1. This was also true for implicit learners, who also showed significant decreasing monotonic trends for both the interleaved training group ( τ = -.272, p < .0001) as well as the blocked training group ( τ = -.336, p = .001) (Fig. (Fig.2 2 ).

Learning curves during training for Explicit and Implicit Learners, Experiment 1 . Significant decreasing trends in all groups reveal learning over 240 trials
Additionally, we conducted one-sample t-tests to determine if learning difference scores differed significantly from zero, which would indicate a change in performance from the beginning to the end of training on Day 1. Participants in the blocked training condition showed faster RTs at the end of training as compared with the beginning of training ( M = -.9585, SD = .7576); t (39) = -8.00, p <.001, d = -1.27. However, using this measure, there was only a weak trend for a significant difference in RT between the first and last sequences in the interleaved condition ( M = -.199, SD = 0.78). t (42) = -1.6657, p = 0.10, d = -.25, despite the significantly decreasing monotonic trends in RT across the session in the interleaved practice condition for both the implicit and explicit learners. To compare learning difference scores between practice conditions, an independent-samples t-test was conducted. As expected, we found that the blocked training group ( M = -.9585, SD = .7576) did show faster learning during training compared with the interleaved group ( M = -.199, SD = 0.78); t (81) = 4.478, p < .001, d = .98. This is consistent with findings in the CI literature in which blocking stimuli in the absence of high contextual interference facilitates fast performance improvements.
A three-way ANOVA was conducted that examined the effect of training condition, testing condition, and learner type (explicit, implicit) on mean RT difference scores. There was a significant main effect of training condition on difference scores, ( F (1, 75) = 39.539 , p < .001, η 2 = 0.274), with less forgetting from Day 1 to Day 2 for participants who had received interleaved training (Fig. (Fig.3). 3 ). Participants who trained in the interleaved condition had a negative difference score, indicating improved performance ( M = -0.313, SD = 0.554). Participants who trained in the blocked condition instead showed a positive difference score, indicating poorer performance from Day 1 to Day 2 ( M = 0.726, SD = 0.938). There was also a significant main effect of testing condition, ( F (1, 75) = 9.538, p = .003, η 2 = 0.066), with greater forgetting for participants who received interleaved testing on Day 2. Participants who received interleaved testing had a mean positive difference score ( M = 0.408, SD = 1.017), while participants who were tested with blocked sequences had a negative mean difference score ( M = -0.049, SD = 0.748).

Mean difference score per condition. Positive difference scores represent increased reaction times (RTs) on Day 2 as compared to Day 1, indicating worse performance. Error bars represent ± SEM
These main effects were qualified by two statistically significant interactions. First, we observed an interaction between the effects of training condition and testing condition on difference scores, F (1,75) = 11.948, p < .001, η 2 = 0.083. An analysis of simple effects showed that testing condition did not significantly affect difference scores when participants were trained in the interleaved condition ( F (1) = 0.027, p = .870). However, testing condition did significantly affect the difference scores when participants were trained in the blocked condition ( F (1) = 20.056, p < .001). Participants who trained with interleaved sequences were able to retain or improve performance regardless of testing condition, while participants who trained with blocked sequences did worse when tested with interleaved sequences.
Additionally, we observed an interaction between the effects of training condition and explicit knowledge on difference scores, F (1, 75) = 4.915, p = .03, η 2 = 0.034. Simple effects analysis showed that learner type did affect difference scores when subjects trained with blocked sequences ( F (1) = 4.529, p = .037), but not when subjects trained with interleaved sequences ( F (1) = 1.082, p = .302). Explicit learners in the blocked training condition had higher difference scores (i.e., more forgetting) than implicit learners, suggesting that explicit learning of the sequences might hinder performance on the delayed test, especially when the sequences were first practiced in a blocked fashion.
In implicit learners, the BI and II groups showed difference scores significantly different than zero. The BI group showed increased RTs on Day 2, indicating forgetting, while the II group showed decreased RTs on Day 2, indicating some consolidation of learning (( t (9) = 3.255, p = .009; t (13) = -3.633, p = .003, respectively). The groups tested with blocked sequences on Day 2 (BB and IB) showed similar RTs across the delay, suggesting good retention of learning across training conditions when sequences were tested in blocks, with no differences between the end of training and the beginning of the test ( t (6) = -.0265, p = .979; t (8) = .119, p = .910, respectively).
In explicit learners, only the BI and IB groups showed difference scores significantly different than zero, in a positive and negative direction, respectively. The BI group showed increased RTs on Day 2 ( t (10) = 5.757, p = .0001), while the IB group demonstrated decreased RTs on Day 2, which reflect faster performance for blocked sequences ( t (11) = -4.849, p = .0005). Difference scores of participants in the II and BB groups did not significantly differ from zero, suggesting good retention from Day 1 ( t (7) = -1.412, p = .201; t (11) = 2.026, p = .068, respectively).
Hours of sleep were assessed; however, participants reported adequate numbers, with little variation ( M = 6.86 , SD = 1.19), and so it was excluded as a covariate.
In this experiment, we used the SRTT to examine the effect of interleaved practice on implicit learning of fine motor sequences after a 1-day delay. We hypothesized that interleaved practice would result in poorer initial performance but superior long-term retention, in line with the CI effect (Shea & Morgan, 1979 ). Consistent with the past findings, we found participants who practiced blocked sequences were much faster than those who practiced interleaved sequences, but they were left unprepared for interleaved sequences at test, especially those with explicit sequence knowledge (Magill & Hall, 1990 ; Shea & Morgan, 1979 ; Wright et al., 2016 ).
We found that interleaved practice may reduce interference from explicit knowledge. An interaction between training condition and learner type (explicit or implicit) revealed that retention was similar for explicit and implicit learners when they practiced interleaved sequences. But for those who practiced blocked sequences, explicit learners were especially impacted, suggesting that explicit knowledge of the sequences may hinder SRTT performance only when sequences are presented in a blocked fashion. This finding is consistent with previous work demonstrating that explicit sequence knowledge can be detrimental to speeded performance in a visuomotor task (Tanaka & Watanabe, 2017 ) as well as on the SRTT (Reber & Squire, 1998 ; but see Willingham et al., 2002 ). It may be that interleaving sequences ameliorates the possible interference that can arise from explicit knowledge, perhaps because practice with high CI encourages general, non-sequence-specific learning that is more immune to intrusions of explicitly learned sequence elements.
The CI effect in motor learning has been reliably demonstrated with explicit sequence learning, but there had been little evidence to suggest that interleaving could benefit sequence learning that occurred without awareness. We found similar results in implicit and explicit learners, in that the CI effect was most pronounced when interleaved sequences were used in the retention test. Even with little to no awareness of any structure within the key presses, those who practiced interleaved sequences showed consolidation of learning when tested with interleaved sequences on Day 2. Those who practiced with blocked sequences were slower when those same sequences were interleaved on Day 2, indicating “forgetting” of sequences that participants were not aware they had learned. These results seem to be in line with the Specificity of Learning hypothesis in that for implicit learners, a consistent practice and testing condition (II) led to better learning than an inconsistent one (BI) (Barnett et al., 1973 ). But those in the IB and BB groups both showed retention, rather than forgetting and consolidation, respectively. This suggests that, similar to other motor studies, our results are more in line with the Variability of Practice hypothesis that emphasizes the role of task variation (e.g., through interleaving) in supporting the learner’s ability to abstract a generalizable schema they can apply to other skills or tasks (Schmidt, 1975 ; Schmidt et al., 1990 ; Shea & Kohl, 1990 ). Blocked practice might result in less flexible learning that is specific to practice conditions, even without conscious awareness of sequences or structure in the task.
In Experiment 1 , interleaved practice led to more flexible retention of practiced sequences in that there was no forgetting for sequences that were presented in either a blocked or interleaved order. In Experiment 2 , we examined whether interleaved practice leads to greater generalization of sequence learning to performance of new sequences. Positive transfer to novel tasks or contexts is a crucial goal in many training situations as one often cannot train on every possible task variation or in every possible context.
Experiment 2
125 right-handed young adults were enrolled in the study (96 female; age 18–30 years, M = 20.6, SD = 1.8). Participants were UCLA undergraduate students and were given course credit for their participation. All participants gave informed consent using an institutionally approved consent form. Participants were excluded if they had a history of neurological or psychiatric disease or if they were taking neuroactive medication that could affect sensory processing, movement, or cognition. All participants reported that they were right-handed, but degree of handedness was not assessed. Participants were excluded if they had an accuracy lower than 80% on either day (Willingham et al., 2000 ). A total of 30 participants were excluded for low accuracy ( n = 16), computer error ( n = 4), and failing to complete the experiment ( n = 10). Our final subject pool consisted of 95 right-handed young adults (72 female, age 18–30 years, M = 20.5, SD = 1.7).
The study design and task is largely the same as Experiment 1 , except that the three sequences presented on Day 2 were novel (e.g., DDD...EEE...FFF for blocked testing or DFEFEDFED….for interleaved testing; Fig. Fig.1). 1 ). Subjects were randomly assigned to one of four training/testing conditions (BB, BI, IB, II). Explicit knowledge of sequences was assessed as in Experiment 1 , except that subjects were asked if they remembered sequences from either Day 1 or Day 2.
Sequence RTs were calculated by summing the eight key press RTs for correct sequences. Data analysis was largely similar to Experiment 1 , except for how the difference score was calculated. To assess transfer, we compared the summed RTs of the first ten presentations of each sequence in the blocked condition or the first 30 sequences in the interleaved condition, and subtracted these from the first ten presentations of each new sequence in the blocked test condition, or the first 30 sequences in the interleaved test condition. Positive transfer would be indicated by faster RTs at the beginning of Day 2 compared to initial performance on Day 1, while negative transfer, or interference, would be indicated by slower RTs. Similar RTs for the beginning of Day 1 and Day 2 would reflect a lack of transfer. We utilized an independent t-test to assess mean items recalled per sequence in both training groups in the explicit knowledge test. We assessed learning during Day 1 training and Day 2 testing using the Mann-Kendall test, a nonparametric test for monotonic trends. Learning difference scores were also calculated by subtracting the RT of the first thirty trials of Day 1 from the RT of the last thirty trials from Day 1.
We also conducted a 2 × 2 between-subjects ANOVA to assess the effect of training and testing conditions on transfer difference scores (beginning of Day 2 – beginning of Day 1). In addition, we conducted an ANCOVA with recall score as the covariate. We also conducted one-sample t-tests to assess which groups (BB, BI, IB, II) had transfer scores that significantly differed from zero.
On Day 1, a Mann-Whitney test indicated that participants who practiced interleaved sequences were significantly less accurate ( M = 91.07, SD = 5.13) than participants who practiced blocked sequences ( M = 94.71, SD = 3.45; U = 1660.00, p < .001). This was also found to be true on Day 2 ( M I = 94.21, SD I = 3.36; M B = 95.72, SD B = 3.07; U = 1455.50, p = .014). Those who were tested on interleaved sequences either received blocked or interleaved training the day before, however training condition did not impact accuracy on Day 2 ( M II = 94.00, SD II = 3.51; M BI = 94.44, SD BI = 3.23; t (48) = 0.47, p = .642). Similarly, training condition did not impact accuracy on Day 2 for those who were tested on blocked sequences ( M BB = 96.30, SD BB = 2.51; M IB = 94.98, SD IB = 3.59; U = 308.50, p = .185).
Explicit recall of sequences was low for both training groups, with no subjects producing more than an average of three sequential items per sequence. On average, subjects recalled fewer than two sequential items per sequence, which is essentially chance recall of sequence elements, indicating that sequence knowledge was substantially implicit. Unlike in Experiment 1 , no participants were excluded from analysis based on substantial explicit sequence knowledge. Though 75.7% of participants reported that they noticed a pattern, only one participant correctly recalled there were six sequences in the entire experiment. There was no significant difference in item recall between participants who trained with interleaved sequences ( M = 1.41 , SD = 1.18) and those who trained with blocked ones ( M = 1.85 , SD = 1.13) ; ( t (96) = -1.8504 , p = .067, d = -0.38). Both groups recalled fewer than two sequential items per sequence on average, demonstrating that all participants learned the sequences implicitly. The reduced amount of explicit sequence knowledge shown by the participants compared to Experiment 1 is likely because Experiment 2 involved six sequences per participant (three sequences on Day 1 followed by three novel sequences on Day 2) and one, not two, sessions of practice on each set of sequences.
We found significant decreasing monotonic trends in RT for both the interleaved training group ( τ = -.431, p < .0001) as well as the blocked training group ( τ = -.39, p < .0001) (Fig. (Fig.4). 4 ). This indicates that both groups showed learning of the sequences over Day 1. On Day 2, significant decreasing monotonic trends were found in all groups, except for BI, indicating that blocked training may hinder new learning of interleaved sequences (Table (Table2; 2 ; Fig. Fig.5 5 ).

Learning curves during training, Experiment 2 . Significant decreasing trends in both groups reveal learning over 240 trials
Results of Mann-Kendall trend test for testing day, Experiment 2
| Condition | Kendall’s Tau | P-value |
|---|---|---|
| II | -.283 | <.0001 |
| BB | -.539 | <.0001 |
| IB | -.531 | <.0001 |
| BI | .0595 | .17 |

Learning curves over Day 2, Experiment 2 . Significant decreasing monotonic trends were found in all groups, except for Blocked-Interleaved, suggesting that blocked training may hinder learning of novel interleaved sequences. Each point is an average of 20 trials
We conducted one-sample t-tests to determine if difference scores from the beginning to the end of training differed significantly from zero, which would indicate a significant change in performance from the beginning to the end of training. Similar to our findings in Experiment 1 , participants in the blocked training condition showed faster RTs at the end of training as compared with the beginning ( M = -1.19; SD = .71); t (48) = -11.74, p < .001; d = -1.68. However, unlike our previous findings, we found that participants in the interleaved training condition did show a significant decrease in RT, demonstrating successful learning as evidenced by a negative difference score ( M = -.31; SD = .64); t (45) = -3.2573, p = .0021, d = -.48. Consistent with our findings in Experiment 1 , we found that the blocked training group ( M = -1.19; SD = .71) showed a greater decrease in RT during training compared to the interleaved group ( M = -.31; SD = .64); t (93) = 6.31, p < .001, d = 1.30.
We next examined transfer learning with a two-way ANOVA of Training condition (Interleaved, Blocked) and Testing condition (Interleaved, Blocked). We found a significant main effect of testing condition, F (1,91) = 9.689, p = .003,η 2 = .092 (Fig. (Fig.6). 6 ). Subjects tested with new sequences in the blocked condition showed greater transfer to the new sequences, as evidenced by a negative difference score ( M = -.608, SD = .588). Subjects tested in the interleaved condition also had a negative difference score, but of a smaller magnitude ( M = -.233, SD = .585). There was a trend for a main effect of training condition ( F (1,91) = 3.469, p = .066,η 2 = .033), with participants receiving interleaved training exhibiting numerically greater transfer to new sequences. The interaction between training and testing conditions was not significant ( F (1,91)=1.225, p =.27,η 2 = .012). However, post hoc Tukey tests showed significant differences between BI and IB groups ( M = 0.599, p = .005) as well as between BI and BB groups ( M = 0.509, p = .015). Those in the BI group showed significantly worse transfer performance than those in the IB and BB groups. This indicates that for blocked practice alone, the type of testing condition affects transfer performance. An ANCOVA controlling for recall score reveals a significant main effect of testing condition ( F (1,90) = 8.895, p = .0004,η 2 = .090). Again, the covariate was not found to be significantly related to performance ( F (1,90) = .057, p = .811,η 2 = .0006).

Transfer learning difference score per condition. Note: Negative difference scores indicate faster reaction times (RTs) on Day 2 as compared to Day 1. All groups except for Blocked/Interleaved demonstrated positive transfer to novel sequences. Error bars represent ± SEM
One-sample t-tests revealed that all groups, except for BI, had transfer scores that differed significantly from zero ( M BI = -.054, SD BI =.511, t (23) = -.516 , p =.611). Thus, II, IB, and BB all showed positive transfer to novel sequences ( M II = -.412, SD II = .428, M IB = -.654, SD IB = .919, M BB = -.563, SD BB = .431; p s <.01). In other words, participants in the blocked training condition showed transfer to the new sequences only when they were presented in a blocked order. Participants who received interleaved training showed significantly faster RTs for the new sequences at the beginning of Day 2 compared with the beginning of Day 1 regardless of test condition.
Like the previous experiment, hours of sleep were assessed but were again excluded as a covariate due to sufficient hours of sleep and little variation ( M = 7.07 , SD = 1.49).
In Experiment 2 , we observed implicit sequence learning and transfer for both blocked and interleaved practice conditions in the SRTT. After interleaved practice, there was substantial transfer to performance of new sequences for both testing conditions. After blocked practice, significant positive transfer occurred if new sequences were blocked, but not if they were presented in an interleaved order. This mirrors our results in Experiment 1 in that blocked practice seemed to be vulnerable to testing condition, while interleaved practice prepared participants for both testing conditions. These results suggest that blocked practice of sequences results in implicit learning that is relatively specific to the mode of practice.
While blocked practice did result in positive transfer to new blocked sequences, these participants performed new interleaved sequences at a similar level to their performance at the onset of initial practice. In contrast, interleaved practice of sequences resulted in implicit learning that facilitated performance of new sequences presented in either a blocked or interleaved fashion. In this way, interleaved practice resulted in learning that was general to the SRTT rather than encapsulated in the practiced sequences, similar to previous findings (Müssgens & Ullén, 2015 ). Additionally, prior research using sequential rule paradigms has found that extensive training with a cognitive task was associated with more errors in a transfer task in which the same rules were used but in a different order, suggesting that sequential expectations about a task can interfere with transfer performance (Woltz et al., 2000 ). Blocked practice may thus be more susceptible to violated sequential expectations, resulting in poorer transfer learning, especially when sequences are interleaved.
This result extends recent findings that prior interleaved practice can result in broader learning benefits that are not specific to the practice session. Interleaved practice is usually associated with poorer acquisition and superior retention, however experience with prior interleaved practice may actually improve acquisition of a novel skill (Hodges et al., 2014 ; Kim et al., 2016 , 2018 ). Individuals who underwent prior interleaved practice showed faster acquisition of a novel task as compared with those with prior blocked practice, thereby mitigating the costs normally associated with high CI during learning. This suggests that experience with high CI may facilitate the rate of learning beyond an isolated practice session, perhaps because the learner is able to apply strategies gleaned from interleaved practice to novel motor tasks. It is possible that prior interleaved practice enabled individuals to generate many different motor programs that could aid future learning of similar motor skills (Kim et al., 2018 ). This may also be reflected in our finding that all groups showed learning during Day 2, except those in the BI group. When novel sequences were interleaved at test, prior blocked practice seemed to hinder participants from learning, while those with prior interleaved practice demonstrated successful learning of novel sequences in both testing conditions.
Summary and concluding discussion
We investigated two hypotheses about the effects of interleaved practice on implicit sequence learning. First, we tested whether interleaved practice of sequences leads to greater retention than blocked practice by examining the effect of practice schedule on sequence RT tested the following day after practice. Next, we tested whether interleaved practice of sequences lead to greater positive transfer to novel sequences than blocked practice by examining the effect of practice schedule on performance of novel sequences the day after practice. We found support for the benefit of interleaved practice on both retention and transfer of implicit sequence learning, indicating that the benefit of interleaved practice does not depend on explicit memory retrieval, but also holds for implicit fine motor learning over a delay. Explicit knowledge of the sequence was detrimental to retention when the sequences were blocked, but not when they were interleaved, suggesting that contextual interference may protect against the interference of explicit knowledge on performance.
The SRTT was used here as it is a relatively simple task that has been used extensively to study implicit learning (Robertson, 2007 ). We used eight-item sequences as these are less likely to be learned explicitly than shorter sequences (Meissner et al., 2016 ; Song et al., 2008 ). Nevertheless, in Experiment 1 , where participants practiced the same three sequences for two days, many participants gained at least some awareness of the sequence, particularly in the BB condition. Because more participants became aware of some elements of the sequences in the blocked conditions, it is possible that the unaware participants in the blocked condition differed in some other systematic way from those who gained awareness. It is unclear why some individuals gained partial explicit knowledge of blocked sequences while others did not – perhaps the large number of practice trials (80 per sequence) were repetitive enough for some observant individuals to notice structure in the task (Willingham, 2001 ). The largest group of implicit learners (n=14) was found in the Interleaved/Interleaved condition (Table (Table1). 1 ). Our results seem consistent with the idea that high CI can encourage implicit learning due to the increased working memory load from frequent task-switching that may make explicit learning more difficult (Rendell et al., 2011 ). In tasks in which explicit knowledge could hinder performance, such as the SRTT (Reber & Squire, 1998 ), CI may facilitate performance by inducing learners to rely on implicit knowledge. Similar results were found by Rendell et al. ( 2011 ), who examined participants’ performance of two gross motor skills of different difficulties while completing a secondary task. Interestingly, participants who practiced a kicking skill with high contextual interference performed exceptionally well under dual-task conditions, suggesting greater implicit learning. The secondary task may require participants to rely on a lesser amount of attentional control and thus these performance gains are due to implicit, rather than explicit, learning. Interleaving tasks may make it more difficult to acquire explicit knowledge, and thus the learner may learn implicitly during acquisition, which in turn maybe more effective for retention and transfer performance. Notably, this result only applied to the more complex skill of kicking, so task difficulty is an important consideration when examining CI (Albaret & Thon, 1998 ; Farrow & Buszard, 2017 ; Guadagnoli & Lee, 2004 ). The Challenge Point Framework predicts that the strength of the CI effect is partially determined by task difficulty; namely that it is more robust with low difficulty tasks (Guadagnoli & Lee, 2004 ). Since Rendell et al. ( 2011 ) only observed the CI effect in the more challenging task, these results seem inconsistent with this framework. In the present study, we used a low difficulty task and were able to observe benefits of CI during practice. Future research should aim to clarify these disparate findings and specifically manipulate task difficulty under varying levels of contextual interference. It is possible that with more complex sequences there would be little or no benefit of CI.
Recent research indicates that frequent error processing in addition to task switching increases cognitive effort and may encourage implicit learning (Broadbent et al., 2017 ). In both experiments presented here we found that those in the interleaved condition were less accurate than those in the blocked condition on Day 1, which may lend some credence to the theory that frequent error processing may occur with high CI (Broadbent et al., 2017 ). However, on Day 2 of Experiment 1 , accuracy did not differ between the two conditions, and we did not observe the costs normally associated with interleaved practice, consistent with past research (Hodges et al., 2014 ) which may reflect a general learning benefit of interleaving that results in improved skill acquisition (Kim et al., 2016 , 2018 ).
In Experiment 2 , we focused on transfer to new sequences. This type of transfer may be conceptually related to playing a new piece of music after extensive practice of a different piece. Implicit learning has been thought to be inflexible and not amenable to transfer (Dienes & Berry, 1997 ; Sanchez et al., 2015 ), though our results suggest that positive transfer of implicit motor sequences may be facilitated by introducing high CI during acquisition. The present results are the first to show a benefit for interleaving in the positive transfer of learned sequences in which the lack of awareness of learning was assessed, indicating that explicit processes may not be required to observe this effect. Past research has found that explicit knowledge can reduce errors but increase RTs, hindering transfer performance overall (Benson et al., 2011 ; Tanaka & Watanabe, 2017 ). However, error processing may be an important component of the learning process as errors allow learners to compare between the actual versus expected outcomes of response selection, which can then inform hypotheses and rules that learners may generate to improve performance (Maxwell et al., 2001 ; Rabbitt, 1966 ). Thus, fewer errors due to explicit knowledge may actually hinder successful long term retention despite superior acquisition performance. As discussed above, this is in line with the idea that interleaving increases working memory load, presumably by task switching and error processing, which encourages an implicit form of learning (Broadbent et al., 2017 ; Rendell et al., 2011 ).
Though promising, our results should be cautiously interpreted due to some limitations. Sample size for both experiments was analogous to similar studies, but the substantial number of explicit learners in Experiment 1 led to a smaller sample size for the implicit learner group. However, we felt that dichotomizing was essential as we were interested in whether the CI effect persists in the absence of explicit knowledge, as implicit learning is an important component of long-term skill retention in both healthy adults (Howard et al., 2004 ; Mazzoni & Krakauer, 2006 ) and for clinical populations with explicit memory deficits (Curran, 1997 ; Gabrieli et al., 1993 ) or those with chronic stroke (Boyd & Winstein, 2004 ; Dimyan & Cohen, 2011 ; Wadden et al., 2017 ). Though prior literature has used similar methods to exclude those with explicit awareness, we acknowledge that this categorization does not address the question of how the effects of interleaving are impacted by increasing levels of awareness of learning. Additionally, some participants who were classified as explicit learners may have only gained explicit knowledge of the sequences towards the end of the retention test, and thus may have learned the sequences implicitly during Day 1. Future research could examine the effects of CI when learning is more clearly implicit in all participants, such as learning with a concurrent task (Grafton et al., 1995 ; Nejati et al., 2008 ) or a more complex probabilistic sequence (Du et al., 2016 ; Song et al., 2007 ).
Fatigue was a potential factor that impacted performance given the large number of trials during training and testing. Extended practice may cause fatigue and participants might fail to show learning despite successful skill acquisition (Eysenck, 1965 ). Performance might improve if fatigue could be mitigated with intervals of rest, but previous work demonstrated that adding breaks to the SRTT did little to change performance in implicit learners (Robertson et al., 2004 ). Additional evidence suggests measuring motor skill learning via RT difference scores is unaffected by fatigue (Heuer et al., 1998 ). However, it is possible that fatigue impacted participants’ motivation or effort, given the number of participants who were excluded based on low accuracy or failing to complete the experiment, despite the SRTT’s relative simplicity. Future research may utilize fewer trials, as implicit sequence transfer may occur even with a shorter learning period (Tanaka & Watanabe, 2015 ).
In sum, the present results add to the literature showing the benefits of interleaved practice on learning and transfer and extend them by providing evidence that this effect is also observed in implicit fine motor sequence learning. Blocked practice of sequences in real world skills, such as repetitively playing single pieces when learning an instrument, may appear to be effective in that performance will improve within the practice session. However, this type of practice may not be as effective for mastering the instrument as interleaved practice of different pieces. Furthermore, these results suggest that patients with deficits in explicit memory can still benefit from high CI to successfully learn and retain novel motor skills.
Open practices statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. Neither of the experiments were preregistered.
Declarations
We have no known conflict of interest to disclose.
This research was supported in part by the National Science Foundation (BCS- 1634157 to B.K.) and UCLA’s Graduate Research Mentorship Fellowship (to J.S.).
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
- Abrahamse, E., Ruitenberg, M., De Kleine, E., & Verwey, W. B. (2013). Control of automated behavior: Insights from the discrete sequence production task. Frontiers in Human Neuroscience , 7 . 10.3389/fnhum.2013.00082 [ PMC free article ] [ PubMed ]
- Albaret J-M, Thon B. Differential effects of task complexity on contextual interference in a drawing task. Acta Psychologica. 1998; 100 (1):9–24. doi: 10.1016/S0001-6918(98)00022-5. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bangert, M., Wiedemann, A., & Jabusch, H.-C. (2014). Effects of variability of practice in music: A pilot study on fast goal-directed movements in pianists. Frontiers in Human Neuroscience , 8 . 10.3389/fnhum.2014.00598 [ PMC free article ] [ PubMed ]
- Barnett ML, Ross D, Schmidt RA, Todd B. Motor Skills Learning and the Specificity of Training Principle. Research Quarterly . American Association for Health, Physical Education and Recreation. 1973; 44 (4):440–447. doi: 10.1080/10671188.1973.10615224. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Battig, W. (1966). Facilitation and Interference. In E. A. Bilodeau (Ed.), Acquisition of Skill (pp. 215–244). Academic Press.
- Benson BL, Anguera JA, Seidler RD. A spatial explicit strategy reduces error but interferes with sensorimotor adaptation. Journal of Neurophysiology. 2011; 105 (6):2843–2851. doi: 10.1152/jn.00002.2011. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bjork, R. A. (1994). Memory and metamemory considerations in the training of human beings. In Metacognition: Knowing about knowing (pp. 185–205). The MIT Press.
- Bjork RA, Kroll JF. Desirable Difficulties in Vocabulary Learning. The American Journal of Psychology. 2015; 128 (2):241–252. doi: 10.5406/amerjpsyc.128.2.0241. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bo J, Seidler RD. Visuospatial Working Memory Capacity Predicts the Organization of Acquired Explicit Motor Sequences. Journal of Neurophysiology. 2009; 101 (6):3116–3125. doi: 10.1152/jn.00006.2009. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bönstrup M, Iturrate I, Thompson R, Cruciani G, Censor N, Cohen LG. A Rapid Form of Offline Consolidation in Skill Learning. Current Biology. 2019; 29 (8):1346–1351. doi: 10.1016/j.cub.2019.02.049. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Boyd LA, Winstein CJ. Providing explicit information disrupts implicit motor learning after basal ganglia stroke. Learning & Memory (Cold Spring Harbor, N.Y.) 2004; 11 (4):388–396. doi: 10.1101/lm.80104. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Brady F. Contextual interference: A meta-analytic study. Perceptual and Motor Skills. 2004; 99 (1):116–126. doi: 10.2466/pms.99.1.116-126. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Broadbent DP, Causer J, Williams AM, Ford PR. The role of error processing in the contextual interference effect during the training of perceptual-cognitive skills. Journal of Experimental Psychology: Human Perception and Performance. 2017; 43 (7):1329. doi: 10.1037/xhp0000375. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Cahill L, McGaugh JL, Weinberger NM. The neurobiology of learning and memory: Some reminders to remember. Trends in Neurosciences. 2001; 24 (10):578–581. doi: 10.1016/S0166-2236(00)01885-3. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Curran T. Higher-Order Associative Learning in Amnesia: Evidence from the Serial Reaction Time Task. Journal of Cognitive Neuroscience. 1997; 9 (4):522–533. doi: 10.1162/jocn.1997.9.4.522. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Dang, K. V., Parvin, D. E., & Ivry, R. B. (2019). Exploring Contextual Interference in Implicit and Explicit Motor Learning. BioRxiv , 644211. 10.1101/644211
- Dienes Z, Berry D. Implicit learning: Below the subjective threshold. Psychonomic Bulletin & Review. 1997; 4 (1):3–23. doi: 10.3758/BF03210769. [ CrossRef ] [ Google Scholar ]
- Dimyan MA, Cohen LG. Neuroplasticity in the context of motor rehabilitation after stroke. Nature Reviews. Neurology. 2011; 7 (2):76–85. doi: 10.1038/nrneurol.2010.200. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Du, Y., Prashad, S., Schoenbrun, I., & Clark, J. E. (2016). Probabilistic Motor Sequence Yields Greater Offline and Less Online Learning than Fixed Sequence. Frontiers in Human Neuroscience , 10 . 10.3389/fnhum.2016.00087 [ PMC free article ] [ PubMed ]
- Eysenck HJ. A three-factor theory of reminiscence. British Journal of Psychology. 1965; 56 (2–3):163–182. doi: 10.1111/j.2044-8295.1965.tb00956.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Farrow, D., & Buszard, T. (2017). Chapter 5—Exploring the applicability of the contextual interference effect in sports practice. In M. R. Wilson, V. Walsh, & B. Parkin (Eds.), Progress in Brain Research (Vol. 234, pp. 69–83). Elsevier. 10.1016/bs.pbr.2017.07.002 [ PubMed ]
- French KE, Rink JE, Werner PH. Effects of contextual interference on retention of three volleyball skills. Perceptual and Motor Skills. 1990; 71 (1):179–186. doi: 10.2466/PMS.71.4.179-186. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Frensch, P. A., & Rünger, D. (2003). Implicit Learning: Current Directions in Psychological Science . https://journals.sagepub.com/doi/10.1111/1467-8721.01213
- Gabrieli JD, Corkin S, Mickel SF, Growdon JH. Intact acquisition and long-term retention of mirror-tracing skill in Alzheimer’s disease and in global amnesia. Behavioral Neuroscience. 1993; 107 (6):899–910. doi: 10.1037/0735-7044.107.6.899. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Goode S, Magill RA. Contextual Interference Effects in Learning Three Badminton Serves. Research Quarterly for Exercise and Sport. 1986; 57 (4):308–314. doi: 10.1080/02701367.1986.10608091. [ CrossRef ] [ Google Scholar ]
- Grafton ST, Hazeltine E, Ivry R. Functional Mapping of Sequence Learning in Normal Humans. Journal of Cognitive Neuroscience. 1995; 7 (4):497–510. doi: 10.1162/jocn.1995.7.4.497. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Guadagnoli MA, Lee TD. Challenge point: A framework for conceptualizing the effects of various practice conditions in motor learning. Journal of Motor Behavior. 2004; 36 (2):212–224. doi: 10.3200/JMBR.36.2.212-224. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Heuer H, Spijkers W, Kiesswetter E, Schmidtke V. Effects of sleep loss, time of day, and extended mental work on implicit and explicit learning of sequences. Journal of Experimental Psychology: Applied. 1998; 4 (2):139–162. doi: 10.1037/1076-898X.4.2.139. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hodges, N. J., Lohse, K. R., Wilson, A., Lim, S. B., & Mulligan, D. (2014). Exploring the Dynamic Nature of Contextual Interference: Previous Experience Affects Current Practice But Not Learning. Journal of Motor Behavior, 46 (6), 455–467. [ PubMed ]
- Howard DV, Howard JH, Japikse K, DiYanni C, Thompson A, Somberg R. Implicit sequence learning: Effects of level of structure, adult age, and extended practice. Psychology and Aging. 2004; 19 (1):79–92. doi: 10.1037/0882-7974.19.1.79. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kang SHK, Pashler H. Learning Painting Styles: Spacing is Advantageous when it Promotes Discriminative Contrast: Spacing promotes contrast. Applied Cognitive Psychology. 2012; 26 (1):97–103. doi: 10.1002/acp.1801. [ CrossRef ] [ Google Scholar ]
- Kantak SS, Winstein CJ. Learning-performance distinction and memory processes for motor skills: A focused review and perspective. Behavioural Brain Research. 2012; 228 (1):219–231. doi: 10.1016/j.bbr.2011.11.028. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Keele SW, Ivry R, Mayr U, Hazeltine E, Heuer H. The cognitive and neural architecture of sequence representation. Psychological Review. 2003; 110 (2):316–339. doi: 10.1037/0033-295X.110.2.316. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kelso JAS, Zanone P-G. Coordination dynamics of learning and transfer across different effector systems. Journal of Experimental Psychology: Human Perception and Performance. 2002; 28 (4):776–797. doi: 10.1037/0096-1523.28.4.776. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kim T, Chen J, Verwey WB, Wright DL. Improving novel motor learning through prior high contextual interference training. Acta Psychologica. 2018; 182 :55–64. doi: 10.1016/j.actpsy.2017.11.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kim T, Rhee J, Wright DL. Allowing time to consolidate knowledge gained through random practice facilitates later novel motor sequence acquisition. Acta Psychologica. 2016; 163 :153–166. doi: 10.1016/j.actpsy.2015.11.012. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kim T, Wright DL. Transcranial Direct Current Stimulation of Supplementary Motor Region Impacts the Effectiveness of Interleaved and Repetitive Practice Schedules for Retention of Motor Skills. Neuroscience. 2020; 435 :58–72. doi: 10.1016/j.neuroscience.2020.03.043. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lee TD, Magill RA, Weeks DJ. Influence of practice schedule on testing schema theory predictions in adults. Journal of Motor Behavior. 1985; 17 (3):283–299. doi: 10.1080/00222895.1985.10735350. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lee, T., & Simon, D. (2004). Contextual interference. In A. Williams & N. Hodges (Eds.), Skill acquisition in sport: Research, theory and practice (pp. 29–44). Routledge.
- Lee TD, Magill RA. The locus of contextual interference in motor-skill acquisition. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1983; 9 (4):730–746. doi: 10.1037/0278-7393.9.4.730. [ CrossRef ] [ Google Scholar ]
- Li Y, Wright DL. An Assessment of the Attention Demands during Random- and Blocked-Practice Schedules. The Quarterly Journal of Experimental Psychology Section A. 2000; 53 (2):591–606. doi: 10.1080/713755890. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lin CH, Fisher BE, Winstein CJ, Wu AD, Gordon J. Contextual Interference Effect: Elaborative Processing or Forgetting—Reconstruction? A Post Hoc Analysis of Transcranial Magnetic Stimulation—Induced Effects on Motor Learning. Journal of Motor Behavior. 2008; 40 (6):578–586. doi: 10.3200/JMBR.40.6.578-586. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lin C-HJ, Knowlton BJ, Chiang M-C, Iacoboni M, Udompholkul P, Wu AD. Brain-behavior correlates of optimizing learning through interleaved practice. NeuroImage. 2011; 56 (3):1758–1772. doi: 10.1016/j.neuroimage.2011.02.066. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lin C-HJ, Wu AD, Udompholkul P, Knowlton BJ. Contextual interference effects in sequence learning for young and older adults. Psychology and Aging. 2010; 25 (4):929–939. doi: 10.1037/a0020196. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Magill RA, Hall KG. A review of the contextual interference effect in motor skill acquisition. Human Movement Science. 1990; 9 (3):241–289. doi: 10.1016/0167-9457(90)90005-X. [ CrossRef ] [ Google Scholar ]
- Maxwell JP, Masters RS, Kerr E, Weedon E. The implicit benefit of learning without errors. The Quarterly Journal of Experimental Psychology. A, Human Experimental Psychology. 2001; 54 (4):1049–1068. doi: 10.1080/713756014. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Mazzoni P, Krakauer JW. An Implicit Plan Overrides an Explicit Strategy during Visuomotor Adaptation. The Journal of Neuroscience. 2006; 26 (14):3642–3645. doi: 10.1523/JNEUROSCI.5317-05.2006. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Meira CM, Jr, Tani G. The contextual interference effect in acquisition of dart-throwing skill tested on a transfer test with extended trials. Perceptual and Motor Skills. 2001; 92 (3,Pt1):910–918. doi: 10.2466/PMS.92.3.910-918. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Meissner, S. N., Keitel, A., Südmeyer, M., & Pollok, B. (2016). Implicit Motor Sequence Learning and Working Memory Performance Changes Across the Adult Life Span. Frontiers in Aging Neuroscience , 8 . 10.3389/fnagi.2016.00089 [ PMC free article ] [ PubMed ]
- Menayo R, Moreno FJ, Sabido R, Fuentes JP, García JA. Simultaneous treatment effects in learning four tennis shots in contextual interference conditions. Perceptual and Motor Skills. 2010; 110 (2):661–673. doi: 10.2466/PMS.110.2.661-673. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Morris CD, Bransford JD, Franks JJ. Levels of processing versus transfer appropriate processing. Journal of Verbal Learning & Verbal Behavior. 1977; 16 (5):519–533. doi: 10.1016/S0022-5371(77)80016-9. [ CrossRef ] [ Google Scholar ]
- Müssgens, D. M., & Ullén, F. (2015). Transfer in Motor Sequence Learning: Effects of Practice Schedule and Sequence Context. Frontiers in Human Neuroscience , 9 . 10.3389/fnhum.2015.00642 [ PMC free article ] [ PubMed ]
- Nader K, Schafe GE, Le Doux JE. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature. 2000; 406 (6797):722–726. doi: 10.1038/35021052. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Nejati V, Farshi MTG, Ashayeri H, Aghdasi MT. Dual task interference in implicit sequence learning by young and old adults. International Journal of Geriatric Psychiatry. 2008; 23 (8):801–804. doi: 10.1002/gps.1976. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Newell, K. (1996). Change in Movement and Skill: Learning, Retention, and Transfer. In M. Latash & M. Turvey (Eds.), Dexterity and Its Development .
- Nissen MJ, Bullemer P. Attentional requirements of learning: Evidence from performance measures. Cognitive Psychology. 1987; 19 (1):1–32. doi: 10.1016/0010-0285(87)90002-8. [ CrossRef ] [ Google Scholar ]
- Obayashi S. Possible mechanism for transfer of motor skill learning: Implication of the cerebellum. The Cerebellum. 2004; 3 (4):204–211. doi: 10.1080/14734220410018977. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Perez CR, Meira CM, Tani G. Does the Contextual Interference Effect Last over Extended Transfer Trials? Perceptual and Motor Skills. 2005; 100 (1):58–60. doi: 10.2466/pms.100.1.58-60. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Rabbitt PM. Errors and error correction in choice-response tasks. Journal of Experimental Psychology. 1966; 71 (2):264–272. doi: 10.1037/h0022853. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Reber PJ, Squire LR. Parallel brain systems for learning with and without awareness. Learning & Memory (Cold Spring Harbor, N.Y.) 1994; 1 (4):217–229. [ PubMed ] [ Google Scholar ]
- Reber PJ, Squire LR. Encapsulation of implicit and explicit memory in sequence learning. Journal of Cognitive Neuroscience. 1998; 10 (2):248–263. doi: 10.1162/089892998562681. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Rendell MA, Masters RSW, Farrow D, Morris T. An implicit basis for the retention benefits of random practice. Journal of Motor Behavior. 2011; 43 (1):1–13. doi: 10.1080/00222895.2010.530304. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Rickard TC, Cai DJ, Rieth CA, Jones J, Ard MC. Sleep does not enhance motor sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2008; 34 (4):834–842. doi: 10.1037/0278-7393.34.4.834. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Robertson EM. The serial reaction time task: Implicit motor skill learning? The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 2007; 27 (38):10073–10075. doi: 10.1523/JNEUROSCI.2747-07.2007. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Robertson EM, Pascual-Leone A, Press DZ. Awareness modifies the skill-learning benefits of sleep. Current Biology: CB. 2004; 14 (3):208–212. doi: 10.1016/j.cub.2004.01.027. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Russell DM, Newell KM. How persistent and general is the contextual interference effect? Research Quarterly for Exercise and Sport. 2007; 78 (4):318–327. doi: 10.1080/02701367.2007.10599429. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sanchez DJ, Yarnik EN, Reber PJ. Quantifying transfer after perceptual-motor sequence learning: How inflexible is implicit learning? Psychological Research. 2015; 79 (2):327–343. doi: 10.1007/s00426-014-0561-9. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Schmidt RA. A schema theory of discrete motor skill learning. Psychological Review. 1975; 82 (4):225–260. doi: 10.1037/h0076770. [ CrossRef ] [ Google Scholar ]
- Schmidt RA, Lange C, Young DE. Optimizing summary knowledge of results for skill learning. Human Movement Science. 1990; 9 (3):325–348. doi: 10.1016/0167-9457(90)90007-Z. [ CrossRef ] [ Google Scholar ]
- Schmidt, R. A., & Lee, T. D. (2005). Motor control and learning: A behavioral emphasis, 4th ed . Human Kinetics.
- Sekiya H. Contextual interference in implicit and explicit motor learning. Perceptual and Motor Skills. 2006; 103 (2):333–343. doi: 10.2466/pms.103.2.333-343. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Shanks DR, St. John MF. Characteristics of dissociable human learning systems. Behavioral and Brain Sciences. 1994; 17 (3):367–395. doi: 10.1017/S0140525X00035032. [ CrossRef ] [ Google Scholar ]
- Shea CH, Kohl RM. Specificity and variability of practice. Research Quarterly for Exercise and Sport. 1990; 61 (2):169–177. doi: 10.1080/02701367.1990.10608671. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Shea, J. B., Hunt, J. P., & Zimny, S. T. (1985). Representational Structure and Strategic Processes for Movement Production. In D. Goodman, R. B. Wilberg, & I. M. Franks (Eds.), Advances in Psychology (Vol. 27, pp. 55–87). North-Holland. 10.1016/S0166-4115(08)62530-3
- Shea, J. B., & Zimny, S. T. (1983). Context Effects in Memory and Learning Movement Information. In R. A. Magill (Ed.), Advances in Psychology (Vol. 12, pp. 345–366). North-Holland. 10.1016/S0166-4115(08)61998-6
- Shea, J. B., & Morgan, R. (1979). Contextual Interference Effects on the Acquisition, Retention, and Transfer of a Motor Skill. Journal of Experimental Psychology: Human Learning and Memory , 5 (2), 179–187.
- Shimizu RE, Wu AD, Knowlton BJ. Cerebellar activation during motor sequence learning is associated with subsequent transfer to new sequences. Behavioral Neuroscience. 2016; 130 (6):572–584. doi: 10.1037/bne0000164. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Shmuelof, L., & Krakauer, J. W. (2014). Recent insights into perceptual and motor skill learning. Frontiers in Human Neuroscience, 8 . 10.3389/fnhum.2014.00683 [ PMC free article ] [ PubMed ]
- Soderstrom NC, Bjork RA. Learning Versus Performance: An Integrative Review. Perspectives on Psychological Science. 2015; 10 (2):176–199. doi: 10.1177/1745691615569000. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Song S, Howard JH, Howard DV. Implicit probabilistic sequence learning is independent of explicit awareness. Learning & Memory. 2007; 14 (3):167–176. doi: 10.1101/lm.437407. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Song S, Howard JH, Howard DV. Perceptual sequence learning in a serial reaction time task. Experimental Brain Research . Experimentelle Hirnforschung. Experimentation Cerebrale. 2008; 189 (2):145–158. doi: 10.1007/s00221-008-1411-z. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Squire LR, Genzel L, Wixted JT, Morris RG. Memory consolidation. Cold Spring Harbor Perspectives in Biology. 2015; 7 (8):a021766. doi: 10.1101/cshperspect.a021766. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Stark-Inbar A, Raza M, Taylor JA, Ivry RB. Individual differences in implicit motor learning: Task specificity in sensorimotor adaptation and sequence learning. Journal of Neurophysiology. 2017; 117 (1):412–428. doi: 10.1152/jn.01141.2015. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tanaka K, Watanabe K. Effects of learning duration on implicit transfer. Experimental Brain Research. 2015; 233 (10):2767–2776. doi: 10.1007/s00221-015-4348-z. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tanaka K, Watanabe K. Explicit instruction of rules interferes with visuomotor skill transfer. Experimental Brain Research. 2017; 235 (6):1689–1700. doi: 10.1007/s00221-017-4933-4. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Travlos AK. Specificity and Variability of Practice, and Contextual Interference in Acquisition and Transfer of an Underhand Volleyball Serve. Perceptual and Motor Skills. 2010; 110 (1):298–312. doi: 10.2466/pms.110.1.298-312. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wadden KP, De Asis K, Mang CS, Neva JL, Peters S, Lakhani B, Boyd LA. Predicting Motor Sequence Learning in Individuals with Chronic Stroke. Neurorehabilitation and Neural Repair. 2017; 31 (1):95–104. doi: 10.1177/1545968316662526. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Walker MP, Brakefield T, Allan Hobson J, Stickgold R. Dissociable stages of human memory consolidation and reconsolidation. Nature. 2003; 425 (6958):616–620. doi: 10.1038/nature01930. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Walker MP, Brakefield T, Morgan A, Hobson JA, Stickgold R. Practice with sleep makes perfect: Sleep-dependent motor skill learning. Neuron. 2002; 35 (1):205–211. doi: 10.1016/S0896-6273(02)00746-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Willingham DB. Becoming aware of motor skill. Trends in Cognitive Sciences. 2001; 5 (5):181–182. doi: 10.1016/S1364-6613(00)01652-1. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Willingham DB, Goedert-Eschmann K. The Relation Between Implicit and Explicit Learning: Evidence for Parallel Development. Psychological Science. 1999; 10 (6):531–534. doi: 10.1111/1467-9280.00201. [ CrossRef ] [ Google Scholar ]
- Willingham DB, Salidis J, Gabrieli JDE. Direct Comparison of Neural Systems Mediating Conscious and Unconscious Skill Learning. Journal of Neurophysiology. 2002; 88 (3):1451–1460. doi: 10.1152/jn.2002.88.3.1451. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Willingham DB, Wells LA, Farrell JM, Stemwedel ME. Implicit motor sequence learning is represented in response locations. Memory & Cognition. 2000; 28 (3):366–375. doi: 10.3758/BF03198552. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Woltz DJ, Gardner MK, Bell BG. Negative transfer errors in sequential cognitive skills: Strong-but-wrong sequence application. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2000; 26 (3):601–625. doi: 10.1037/0278-7393.26.3.601. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wright D, Verwey W, Buchanen J, Chen J, Rhee J, Immink M. Consolidating behavioral and neurophysiologic findings to explain the influence of contextual interference during motor sequence learning. Psychonomic Bulletin & Review. 2016; 23 (1):1–21. doi: 10.3758/s13423-015-0887-3. [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wymbs NF, Grafton ST. Neural Substrates of Practice Structure That Support Future Off-Line Learning. Journal of Neurophysiology. 2009; 102 (4):2462–2476. doi: 10.1152/jn.00315.2009. [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
The Contextual Interference Effect Explained

As athletes, coaches and sport scientists we are interested in any factors that can speed up skill acquisition. Contextual interference is one factor that has been shown to accelerate motor learning . In this article we’ll explain contextual interference, discuss how you can apply it in practice and explore the theory behind this effect.
What is contextual interference?
Contextual interference involves mixing up your practice structure, either by adding different tasks and/or adding practice variability when learning a skill. The contextual interference effect shows that this interference results in worse practice performance, but a greater ability to retain and transfer skill at a later date.
This effect was first seen during a word learning task in the 1960s, and was further demonstrated when learning movements in the late 1970s. You can find a full historic review of the contextual interference effect here .
Why is the contextual interference effect useful?
This effect is counterintuitive, as we tend to associate better practice performance with athletes learning more. Whereas, the contextual interference effect tells us that this may not always be the case. By adding in more practice variability, we challenge the athlete, they make more errors, but this seems to create a longer lasting change in performance (increased learning).
What is contextual interferences & practice variability?
Here is where both students and coaches can get confused. There are two continuums along which we can add practice variability and contextual interference (CI) into our practice. Historically, this small detail hasn’t been explained very well in the literature.
The classic CI effect involves adding in new tasks to practice. We call this scale blocked to random practice. In the example below, we’ll use a footballer practicing her throw ins.
Blocked practice (bottom left corner) would be her completing 20 throw ins in a row, this practice would be deemed to have no contextual interference. As we move up the left hand side we are adding in different tasks. An example would be 20 throw ins, 20 passes and 20 sprints all carried out in a random order (top left corner). This would be called random practice and be considered to have higher levels of contextual interference.

The second scale sees us move left to right across our grid, from constant to varied practice. If we are being pedantic we could call this adding practice variability , but CI studies seem to use this continuum too for exploring the CI effect.
In the bottom left corner our footballer would be practicing all her throw ins to the same target, we call this constant practice . As we move to the right we can add variability into the task, one example would be to vary the distance of her target, but we could vary direction too. We would call this varied practice .
Using this grid you can start to see that we can describe practice using both these scales:
- Blocked – constant practice (bottom left)
- Blocked – random practice (top left)
- Varied – constant practice (bottom right)
- Varied – random practice (top right)
It is important to note that, in reality, these are sliding scales, rather than four separate types of practice. However, understanding how to manipulate practice along these two scales can be very useful as a coach and athlete.
Both, adding in random practice and adding in varied practice have been used to explore contextual interference effect in sport. As the grid shows, the bottom left corner has the least CI, then the more we move up and right, the more contextual interference we have within our practice structure.
What is optimal for learning?
It should be noted that the majority of the research has taken place with beginners learning simple, closed skills and despite results being statistically significant, some researchers have questioned the effect size and usefulness of such findings.
Despite the disagreement, there is consistent evidence that contextual interference improves the rate at which beginners learn tasks. One of the most useful recent papers showed that systematically increasing contextual interference with beginners outperformed both blocked and varied practice.
This finding suggests that we should start coaching beginners using constant-blocked practice, then slowly introduce more varied or random practice as they progress.
What causes the contextual interference effect?
Historically, the contextual interference effect has been explained from a cognitive standpoint. Every sports task requires a level of understanding and planning. By mixing up tasks during practice the athlete has to plan, forget and re-plan each time, this is not required when carrying out the same task over and over again (constant-blocked practice). This theory is called the forgetting-reconstruction hypothesis .
A similar, but slightly differing theory is called the elaboration hypothesis . Whereby mixing up tasks creates more distinctive memories between reps and induces better retention in our long term working memory. Some studies have attempted to confirm these theories using electrophysiology data, but there is some way to go before we properly understand the neural workings behind this behavioural effect.
Check out this chapter for more around theoretical underpinnings and contemporary issues surrounding the contextual interference effect.
Summarising contextual interference
Based on the evidence to date, it appears practice variability can result in slightly worse practice performance, but better retention and transfer of sporting skill. There is further evidence to indicate systematically increasing contextual interference could be the best approach when coaching beginners.
The size of the effect is still under debate and there is still limited data on this effect within elite performers. Further research is also needed to understand the neural working that underpins the contextual interference effect.
For more insights into practice structure check out this article on massed vs distributed practice .
Further reading
- Brady (2008) The contextual interference effect and sport skills
- Farrow & Buszard (2017) Exploring the applicability of the contextual interference effect in sports practice
- Lin et al. (2008) Contextual interference effect: elaborative processing or forgetting-reconstruction? A post hoc analysis of transcranial magnetic stimulation-induced effects on motor learning
- Magill & Hall (1990) A review of the contextual interference effect in motor skill acquisition
- Porter & Magill (2010) Systematically increasing contextual interference is beneficial for learning sport skills
Page Reference
If you quote information from this page in your work, then the reference for this page is:
- Shaw, W (2021). Contextual Interference Effect Explained . Available from: https://sportscienceinsider.com/contextual-interference-effect. [Accessed dd/mm/yyyy].
How useful was this post?
Click on a star to rate it!
Average rating 4.4 / 5. Vote count: 8
No votes so far! Be the first to rate this post.
We are sorry that this post was not useful for you!
Let us improve this post!
Tell us how we can improve this post?

Will Shaw PhD, MSc
Will is a sport scientist and golf professional who specialises in motor control and motor learning. Will lecturers part-time in motor control and biomechanics, runs Golf Insider UK and consults elite athletes who are interested in optimising their training and performance.
- Knowledge of Results vs Knowledge of Performance
- Skill Classification Continuums – Learn the Basics
- Motor Learning Explained
- Performance Coaching & Skill Acquisition in Elite Golf
Action plan reconstruction during every attempt in random practice.

Context in source publication
Similar publications.

- EDUC PSYCHOL REV

- Flavio Junior Guidotti

- Julius Baba Apidogo

- Shih-Wei Huang

- Kuei‐Yen Tsai

- J MOTOR BEHAV

- Mariele crociani

- Marcella De Castro Velten
- Gylton Brandão
- Willy Candra

- Matteo Zago

- J NEUROENG REHABIL
- Chang Yoon Baek
- Hyeong Dong Kim
- Dong Yup Yoo
- Kyoung Yee Kang
- Jang Woo Lee
- Recruit researchers
- Join for free
- Login Email Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google Welcome back! Please log in. Email · Hint Tip: Most researchers use their institutional email address as their ResearchGate login Password Forgot password? Keep me logged in Log in or Continue with Google No account? Sign up
An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
- My Bibliography
- Collections
- Citation manager
Save citation to file
Email citation, add to collections.
- Create a new collection
- Add to an existing collection
Add to My Bibliography
Your saved search, create a file for external citation management software, your rss feed.
- Search in PubMed
- Search in NLM Catalog
- Add to Search
Contextual interference effect: elaborative processing or forgetting-reconstruction? A post hoc analysis of transcranial magnetic stimulation-induced effects on motor learning
Affiliation.
- 1 Division of Biokinesiology and Physical Therapy, School of Dentistry, University of Southern California, Los Angeles 90033, USA.
- PMID: 18980910
- DOI: 10.3200/JMBR.40.6.578-586
The elaborative-processing and forgetting-reconstruction hypotheses are the 2 principal explanations for the contextual interference (CI) effect. The present authors' purpose was to identify which of these 2 hypotheses better accounts for the CI effect. They synchronized single transcranial magnetic stimulation (TMS) pulses to each intertrial interval to modulate information processing during Blocked and Random Practice conditions. Participants practiced 3 arm tasks with either a Blocked or Random Practice order. The 3 stimulation conditions (No TMS, TMS, Sham TMS) by 2-practice order (Blocked, Random) between-participant design resulted in 6 experimental groups. Without TMS, motor learning increased under Random Practice. With TMS, this learning benefit diminished. These results support the elaborative-processing hypothesis by showing that perturbing information processing, evoked by Random Practice, deteriorates the learning benefit. Unlike the prediction of the forgetting-reconstruction hypothesis, adding perturbation during Blocked Practice did not significantly enhance motor learning.
PubMed Disclaimer
Similar articles
- Unifying practice schedules in the timescales of motor learning and performance. Verhoeven FM, Newell KM. Verhoeven FM, et al. Hum Mov Sci. 2018 Jun;59:153-169. doi: 10.1016/j.humov.2018.04.004. Epub 2018 Apr 21. Hum Mov Sci. 2018. PMID: 29684760 Review.
- Neural correlates of the contextual interference effect in motor learning: a transcranial magnetic stimulation investigation. Lin CH, Winstein CJ, Fisher BE, Wu AD. Lin CH, et al. J Mot Behav. 2010 Jul-Aug;42(4):223-32. doi: 10.1080/00222895.2010.492720. J Mot Behav. 2010. PMID: 20570818 Clinical Trial.
- Effect of task practice order on motor skill learning in adults with Parkinson disease: a pilot study. Lin CH, Sullivan KJ, Wu AD, Kantak S, Winstein CJ. Lin CH, et al. Phys Ther. 2007 Sep;87(9):1120-31. doi: 10.2522/ptj.20060228. Epub 2007 Jul 3. Phys Ther. 2007. PMID: 17609332 Clinical Trial.
- The role of the primary motor cortex during skill acquisition on a two-degrees-of-freedom movement task. Shemmell J, Riek S, Tresilian JR, Carson RG. Shemmell J, et al. J Mot Behav. 2007 Jan;39(1):29-39. doi: 10.3200/JMBR.39.1.29-39. J Mot Behav. 2007. PMID: 17251169
- Effects of attentional focus, self-control, and dyad training on motor learning: implications for physical rehabilitation. McNevin NH, Wulf G, Carlson C. McNevin NH, et al. Phys Ther. 2000 Apr;80(4):373-85. doi: 10.1093/ptj/80.4.373. Phys Ther. 2000. PMID: 10758522 Review.
- Influence of Heat Exposure on Motor Control Performance and Learning as Well as Physiological Responses to Visuomotor Accuracy Tracking Task. Aoki M, Yamazaki Y, Otsuka J, Okamoto Y, Takada S, Shirai N, Fujimoto T, Ochi G, Yamashiro K, Sato D, Amano T. Aoki M, et al. Int J Environ Res Public Health. 2022 Sep 28;19(19):12328. doi: 10.3390/ijerph191912328. Int J Environ Res Public Health. 2022. PMID: 36231630 Free PMC article.
- Interleaved practice benefits implicit sequence learning and transfer. Schorn JM, Knowlton BJ. Schorn JM, et al. Mem Cognit. 2021 Oct;49(7):1436-1452. doi: 10.3758/s13421-021-01168-z. Epub 2021 Apr 1. Mem Cognit. 2021. PMID: 33796961 Free PMC article.
- Examining Different Motor Learning Paradigms for Improving Balance Recovery Abilities Among Older Adults, Random vs. Block Training-Study Protocol of a Randomized Non-inferiority Controlled Trial. Nachmani H, Paran I, Salti M, Shelef I, Melzer I. Nachmani H, et al. Front Hum Neurosci. 2021 Feb 25;15:624492. doi: 10.3389/fnhum.2021.624492. eCollection 2021. Front Hum Neurosci. 2021. PMID: 33716695 Free PMC article.
- Neural Mechanisms of the Contextual Interference Effect and Parameter Similarity on Motor Learning in Older Adults: An EEG Study. Beik M, Taheri H, Saberi Kakhki A, Ghoshuni M. Beik M, et al. Front Aging Neurosci. 2020 Jun 12;12:173. doi: 10.3389/fnagi.2020.00173. eCollection 2020. Front Aging Neurosci. 2020. PMID: 32595488 Free PMC article.
- Challenge to Promote Change: The Neural Basis of the Contextual Interference Effect in Young and Older Adults. Pauwels L, Chalavi S, Gooijers J, Maes C, Albouy G, Sunaert S, Swinnen SP. Pauwels L, et al. J Neurosci. 2018 Mar 28;38(13):3333-3345. doi: 10.1523/JNEUROSCI.2640-17.2018. Epub 2018 Feb 26. J Neurosci. 2018. PMID: 29483284 Free PMC article.
Publication types
- Search in MeSH
Related information
- PubChem Compound
- PubChem Substance
LinkOut - more resources
Full text sources.
- Taylor & Francis
- Citation Manager
NCBI Literature Resources
MeSH PMC Bookshelf Disclaimer
The PubMed wordmark and PubMed logo are registered trademarks of the U.S. Department of Health and Human Services (HHS). Unauthorized use of these marks is strictly prohibited.
Advertisement
Interleaved practice benefits implicit sequence learning and transfer
- Open access
- Published: 01 April 2021
- Volume 49 , pages 1436–1452, ( 2021 )
Cite this article
You have full access to this open access article

- Julia M. Schorn 1 &
- Barbara J. Knowlton 1
5129 Accesses
10 Citations
7 Altmetric
Explore all metrics
Compared to blocked practice, interleaved practice of different tasks leads to superior long-term retention despite poorer initial acquisition performance. This phenomenon, the contextual interference effect, is well documented in various domains but it is not yet clear if it persists in the absence of explicit knowledge in terms of fine motor sequence learning. Additionally, while there is some evidence that interleaved practice leads to improved transfer of learning to similar actions, transfer of implicit motor sequence learning has not been explored. The present studies used a serial reaction time task where participants practiced three different eight-item sequences that were either interleaved or blocked on Day 1 (training) and Day 2 (testing). In Experiment 1 , the retention of the three training sequences was tested on Day 2 and in Experiment 2 , three novel sequences were performed on Day 2 to measure transfer. We assessed whether subjects were aware of the sequences to determine whether the benefit of interleaved practice extends to implicitly learned sequences. Even for participants who reported no awareness of the sequences, interleaving led to a benefit for both retention and transfer compared to participants who practiced blocked sequences. Those who trained with blocked sequences were left unprepared for interleaved sequences at test, while those who trained with interleaved sequences were unaffected by testing condition, revealing that learning resulting from blocked practice may be less flexible and more vulnerable to testing conditions. These results indicate that the benefit of interleaved practice extends to implicit motor sequence learning and transfer.
Similar content being viewed by others
Optimizing performance through intrinsic motivation and attention for learning: the optimal theory of motor learning.

Can we enhance working memory? Bias and effectiveness in cognitive training studies
The reliability paradox: why robust cognitive tasks do not produce reliable individual differences.
Avoid common mistakes on your manuscript.
Introduction
From the moment we wake up to the moment we fall asleep, we perform many skills that we have learned over time, like brushing teeth, typing, driving a car, or playing a musical instrument. Complex and simple skills alike rely on motor dexterity, sequence learning, perceptual acuity, and both explicit and implicit learning (Shmuelof & Krakauer, 2014 ). Procedural skill learning is a persistent and crucial part of the human experience, so determining an optimal practice schedule is essential as we are heavily dependent on our ability to learn new skills throughout our lifetime. Practice schedules that introduce high contextual interference (CI) by interleaving, or randomizing, tasks hinder initial performance but aid in long-term skill retention. While the CI effect is robust, it has yet to be reliably demonstrated in implicit motor sequence learning and transfer. Determining if the CI effect persists in the absence of explicit knowledge is essential in order to design effective practice schedules, especially for clinical populations who demonstrate impaired explicit memory but preserved implicit memory. For example, patients with amnesia show intact motor sequence learning but impaired declarative memory (Reber & Squire, 1994 ). Optimizing practice schedules to enhance long-term retention and transfer of learning will benefit lives across a broad spectrum of settings.
Decades of cognitive psychology research have demonstrated that CI through interleaved, or random, practice (compared to blocked practice) is a way to effectively acquire skills that can be retained in the long-term (Battig, 1966 ; Magill & Hall, 1990 ; Shea & Morgan, 1979 ). Interleaving tasks or stimuli can be thought of as a “desirable difficulty” as it hinders initial performance but results in superior long-term retention and transfer performance (Bjork, 1994 ). Blocked practice, on the other hand, facilitates acquisition performance because it requires the learner repeatedly performs a single task before moving on to the next one. In one of the earliest demonstrations of the benefits of CI in motor skill learning, subjects practiced three sequential arm movement tasks, which were either blocked together or intermixed in an interleaved condition (Shea & Morgan, 1979 ). While performance was worse in the interleaved condition during practice, subjects showed better retention and transfer to similar tasks. Blocked practice usually facilitates faster acquisition but results in poorer long-term retention. This differential effect exemplifies the difference between learning and performance, a critical distinction (for a review, see Soderstrom & Bjork, 2015 ). For example, a pianist may quickly master a difficult passage by practicing it repeatedly only to forget all progress the next day, because conditions that enhance performance may not enhance learning.
At first glance, the CI effect seems to be at odds with the Specificity of Learning hypothesis, which posits that learning is most optimal when the practice conditions during the acquitision phase are the same as those during the testing phase (Barnett et al., 1973 ). Some findings in the motor learning literature suggest there is partial support for this hypothesis, though the CI effect is more robust (Shea & Kohl, 1990 ; Travlos, 2010 ). Thus, interleaved practice shows benefits even when tasks are blocked at test.
Theorized mechanisms of the contextual interference effect
Though the benefits of contextual interference are well documented, the mechanism by which interleaving facilitates this enhanced retention is still not completely understood. Two general hypotheses currently stand out. The forgetting-reconstructive hypothesis (or action plan reconstruction hypothesis) posits that interleaving is beneficial because it requires each task set to be frequently retrieved, while during blocked practice the task set remains in working memory for the entire task duration (Lee & Magill, 1983 ; Lee et al., 1985 ). When practicing interleaved motor sequences, a pattern must be learned and then immediately “dumped” from working memory in order to prepare for subsequent trials (Lee & Simon, 2004 ). In each trial, the learner must retrieve a motor pattern into working memory or construct one from scratch. Conversely, blocked motor sequences can remain in working memory for multiple trials without needing to be updated. While frequently forgetting and retrieving stimuli may initially hinder performance, it also allows the learner to practice reconstructing motor patterns, which is beneficial for long-term retention. This hypothesis may partially explain the dissociation between learning and performance often seen in the CI effect (for a review, see Kantak & Winstein, 2012 ).
On the other hand, the elaboration-distinctiveness hypothesis (or discriminative-contrast hypothesis) posits that interleaved practice facilitates organizational and item-specific processing, so that subjects frequently compare different stimuli for more durable encoding (Shea et al., 1985 ; Shea & Zimny, 1983 ). When tasks or stimuli are interleaved, differences between them may be more easily discerned as compared to when they are blocked.
Though traditionally pitted against one another, these two theories are not mutually exclusive and may explain different components of the CI phenomenon (Lin et al., 2008 ). In an fMRI study, interleaved practice of a motor sequence task compared to blocked practice resulted in increased activation in sensorimotor regions, followed by a decreased activation in similar regions during a delayed retention test (Lin et al., 2011 ). The increased activation during encoding of interleaved sequences was interpreted as representing additional motor program reloading. Since there was increased retrieval processing during encoding, retrieval at test required less activation in these regions. This view lends credence to the forgetting-reconstructive hypothesis.
However, it appears that interleaving may also offer greater benefits than simply requiring more retrieval. In a study where participants learned to identify paintings by artist in either a blocked or interleaved fashion to develop a concept of different artists’ styles, participants in the interleaved condition showed better learning than those who studied the paintings in a blocked but spaced schedule (Kang & Pashler, 2012 ). As temporal spacing of study items required explicit retrieval of the artist’s names, inferior performance in the blocked condition indicated that retrieval practice alone cannot account for the benefits of interleaving. Participants had to abstract information across paintings to learn artists’ styles that could be used to classify new paintings, so the contrasts and comparisons made at encoding in the interleaved condition may have been important for generalization, supporting the elaboration-distinctiveness view.
Neither of these theories specifically account for implicit processes important for motor skill learning, and research on the benefits of interleaving has largely ignored the interplay between implicit and explicit learning (Bjork & Kroll, 2015 ; Shanks & St. John, 1994 ). In light of this, an alternate theory has emerged that proposes that high CI and increased task switching overloads an individual’s working memory capacity, preventing them from gaining explicit task-relevant knowledge (Rendell et al., 2011 ) and enabling greater implicit learning. Previous research showing increased levels of cognitive activity with interleaved practice as compared to blocked practice lends some support to this theory (Li & Wright, 2000 ). Recent findings extend this idea, suggesting that high cognitive effort seen in interleaved practice may be partially due to increased error processing as well as task-switching (Broadbent et al., 2017 ). Following an error, greater cognitive activity could be attributed to an individual updating and correcting a rule as well as retrieving information for the upcoming task. However, evidence supporting this hypothesis is sparse and relegated to perceptual-cognitive skills. It is also unclear whether CI simply enables more acquisition of implicit knowledge, or if it leads to implicit knowledge that is better retained and better able to support transfer to similar tasks. In the present study, implicit learning under blocked and interleaved conditions will be compared directly.
The contextual interference effect and implicit motor sequence learning
Research concerning the CI effect and implicit motor learning has largely focused on gross motor skills like those used when playing sports (French et al., 1990 ; Goode & Magill, 1986 ; Menayo et al., 2010 ). Furthermore, research investigating the CI effect in fine motor sequence learning has almost exclusively focused on explicit memory (Wright et al., 2016 ). Though previous research has explored the effect of CI in implicit motor learning, few studies specifically investigate fine motor sequence learning over a substantial delay (Dang et al., 2019 ; Sekiya, 2006 ) and is thus a primary aim of this paper. One such study examined the CI effect in a pursuit-tracking task (Sekiya, 2006 ). The experimenters told participants in the explicit group the presentation order and number of patterns in the task, while those in the implicit group received no instruction. Interestingly, they failed to replicate the CI effect and found no differences between implicit and explicit learners. One possibility for this finding is the relatively high CI in the Blocked group may have reduced differences between the practice conditions. Blocked segments were intermixed with random segments so that participants did not become aware of the repetition. Thus, it is not yet clear if implicit and explicit learning are similarly affected by CI during practice.
Implicit motor sequence transfer
Positive transfer to novel tasks or contexts is a crucial goal in many training situations as one often cannot train on every possible task variation or in every possible context. It is suggested that when positive transfer occurs, a memory representation of the skill has been created that is more general than a representation that could only support the practiced sequence. Positive transfer would indicate that participants learned not only the practiced sequences but also generalized knowledge that benefits the performance of new sequences. Learning may also be sequence-specific, with performance of new sequences similar to performance at the beginning of practice on the original sequences. However, learning could also result in negative transfer with performance of new sequences impaired due to interference (Obayashi, 2004 ). In this case, learning may be sequence-specific, and this sequence knowledge may impair the ability to perform similar sequences.
Though the benefits of interleaving on retention are well-studied, there is currently less evidence that interleaving can also lead to improved transfer to similar actions (Bangert et al., 2014 ; Brady, 2004 ; Meira & Tani, 2001 ; Russell & Newell, 2007 ; Schmidt & Lee, 2005 ). Transfer in the skill learning domain has been extensively studied, including transfer of learning from one effector to another (Kelso & Zanone, 2002 ), such as right hand to left hand, or scaling, such as performing a skilled action at a different rate or using greater force (Newell, 1996 ). However, there are limited transfer studies on fine motor skills, and none that specifically consider implicit sequence learning over a long delay. Müssgens and Ullén ( 2015 ) showed that interleaved sequences, as compared to blocked, led to better transfer to a new sequence on an immediate test, but it is unlikely that these sequences were implicitly learned as a majority of subjects reported some sequence awareness. Additionally, the immediate test made it likely that interference occurred on the test with the sequences that had just been practiced. Similarly, interleaving may reduce negative transfer of motor sequences when new sequences are performed (Shimizu et al., 2016 ). However, transfer was tested at the end of the experiment, and not after a one day delay when the effects of contextual interference are most apparent. This delay is also crucial for observing “offline gains” in which motor memory is stabilized and improved in the absence of practice and is influenced by sleep (Nader et al., 2000 ; Walker et al., 2002 ). It was also unclear whether sequence knowledge was primarily explicit or implicit in Shimizu et al. ( 2016 ).
Other studies concerning the CI effect in novel motor skill learning found that prior experience with interleaved practice improved new motor task acquisition, however, delayed retention was unaffected by practice schedule (Hodges et al., 2014 ; Kim et al., 2016 , 2018 ). Hodges et al. ( 2014 ) found that random practice experience, compared with blocked practice, led to better acquisition of three novel motor skills after a one-day delay and mitigated the low accuracy cost usually associated with random practice, but ultimately did not affect retention. Using a similar paradigm, Kim et al. ( 2016 ) found that interleaved practice of a motor skill was beneficial for novel skill acquisition, but afforded no retention benefits after a delay. However, interleaved practice in this experiment was not truly random and was more analogous to a blocked practice schedule, as participants practiced 5–15 repetitions of the same motor sequence in a 30-s trial. Kim et al. ( 2018 ) sought to address this limitation by using the discrete sequence production (DSP) task to induce high levels of contextual interference and found that acquisition of a novel task was better for individuals with prior interleaved rather than blocked practice, and this benefit remained after a significant delay. Notably, awareness for the sequences was not probed in any of these experiments and sequence learning was presumed to be explicit. Though participants may not always possess explicit, in-depth knowledge of the task, the DSP task is considered to be an explicit sequence learning paradigm and participants are often informed that they are performing fixed motor sequences (Abrahamse et al., 2013 ; Bo & Seidler, 2009 ). In light of this, our second experiment aims to examine if contextual interference can aid in positive transfer of novel motor sequences when learning is implicit. Demonstrating the CI effect in transfer of sequence learning to different (but similar) sequences would show that this effect persists despite a high degree of potential interference, similar to real-life learning.
The current experiment
The proposed study aimed to add to the body of research on contextual interference by examining the effect of interleaved practice on implicit learning of sequences in a serial reaction time task (SRTT). In this task, participants can acquire knowledge of a sequence of locations as shown by more rapid responses to locations presented in sequence compared to those presented in a random order (Nissen & Bullemer, 1987 ). Participants can show sequence-specific improvement without conscious awareness that the locations appeared in any sequence. Additionally, participants might show non-sequence-specific learning, in that learning and performance of new sequences may be faster than initial performance. Though the SRTT typically examines learning that is specific to the practiced sequence, we argue that learning that generalizes to new sequences may be more relevant to real-world skill learning. In both experiments, training and testing comprised of blocked and interleaved practice conditions, allowing us to examine transfer-appropriate processing (TAP) effects (Morris et al., 1977 ). TAP effects are related to the Specificity of Learning hypothesis which states that performance is optimal when the context during testing resembles the context during training (Barnett et al., 1973 ). There is some support of this hypothesis in the motor learning literature; however, the CI effect is more robust and does not always align with this principle, in that test performance in either condition is often better when preceded by interleaved practice (Shea & Kohl, 1990 ; Travlos, 2010 ).
General method
We examined the effect of practice schedule on motor sequence learning and transfer by either blocking or interleaving the presentation of three different eight-item motor sequences in a two-day experiment utilizing the SRTT. Both experiments thus had four groups, based on condition, for Day 1 (Train) and Day 2 (Test): Blocked training/Blocked test (BB), Interleaved training /Interleaved test (II), Blocked training/Interleaved test (BI), and Interleaved training/Blocked test (IB). At the end of the experiment, explicit knowledge of the sequences was probed with a questionnaire.
Behavioral task
The SRTT has frequently been used to measure implicit learning (Keele et al., 2003 ; Robertson, 2007 ). In this simple task, the participants are asked to respond to cued locations using keypresses. The participant is instructed to respond to the successive locations as quickly and accurately as possible and is not informed that there is a structure governing the order of the appearance of the cued locations. Despite the lack of awareness of the structure, reaction time (RT) is faster for sequences that were practiced compared with RT for a random presentation of cued locations. RT is the primary dependent measure because error rates are generally low and accuracy is not a useful measure of motor learning in this task. This task may share many features of real-world skills that engage fine motor circuits in which movement components must be produced in a specific sequence, such as typing or playing a musical instrument.
Study design
We utilized a between-subjects design with the SRTT. Subjects sat with four fingers of the right hand on four keys on a keyboard (U, I, O, P) that corresponded to the four outlined, unfilled circles on a blue computer screen in a darkened room. One of the circles turned white to act as a cue to press the corresponding key (i.e., the first circle on the screen corresponds with “U”). After the button press, another circle turned white and the first circle reverted to being unfilled. A tone sounded if a subject failed to press a button or if a subject made an incorrect button press. RT and accuracy were recorded for each key press. No baseline performance was measured, since this task is simple and baseline performance does not reliably capture individual differences in learning (Stark-Inbar et al., 2017 ).
Participants practiced three different eight-item sequences that were either interleaved or blocked on Day 1 (training) and Day 2 (testing) (Fig. 1 ). To control for specific item effects, the sequences were randomized so that no two participants had the same ones. Each sequence contained each letter (U, I, O, P) twice. Critically, participants were not told that there were sequences, only to respond to each cue as quickly and as accurately as possible. Participants were randomized into the Interleaved or Blocked training groups and were tested in either a Blocked or an Interleaved condition, counterbalanced across training conditions. Participants were not told which condition they were assigned to on either day. In the blocked condition, participants received 80 repeated presentations of each sequence (i.e., AAA…BBB...CCC). In the interleaved condition, they received three sequences interleaved for a total of 240 trials (i.e., ACBABCBAC....). This number of trials is comparable to the training procedure used in our previous work (Lin et al., 2010 ).

Sequences and study design. ( A ) Six example sequences. In Experiment 1 , participants learned three sequences; in Experiment 2 , participants learned six sequences – three on Day 1 and three novel sequences on Day 2. ( B ) Study design for both experiments. Day 1 is the same for both while Day 2 is separated out by experiment. Each number represents an eight-item sequence as shown in ( A )
Day 2 was the same as Day 1, participants were randomly assigned to either the blocked or the interleaved condition. In Experiment 1 , the sequences presented on Day 2 to each participant were the same as those presented on Day 1. In Experiment 2 , the sequences presented on Day 2 were novel, so participants practiced three sequences on Day 1 and three different sequences on Day 2, for a total of 6 unique sequences. Each day in each experiment contained the same number of trials.
To assess whether subjects had acquired explicit knowledge of the sequences, a free-recall questionnaire based on past research was administered after the second session (Robertson et al., 2004 ; Willingham & Goedert-Eschmann, 1999 ). Though free recall may also reflect a degree of implicit memory, that is also true of most explicit awareness tests (Shanks & St. John, 1994 ). Though there is no consensus on the best way to measure awareness in this task (Robertson, 2007 ), free recall, rather than recognition, was assessed as it can be argued that it is the most appropriate measure of explicit memory (Frensch & Rünger, 2003 ). The test was comprised of the three following questions, which prompted the participants to recall the sequences:
Did you notice any pattern(s)?
How many sequences where there?
Please type in all sequences (using the same keys) and hit Enter after each sequence.
Participants were also asked how many hours of sleep they got in between Day 1 and Day 2. Sleep has been shown to be critical for offline gains in implicit learning, however, rapid consolidation of a motor skill can occur within hours or even seconds after practice (Bönstrup et al., 2019 ; Kim & Wright, 2020 ; Squire et al., 2015 ; Walker et al., 2002 ). We chose a 24-h interval between training and testing to measure long-term retention as this is sufficient to observe consolidation; subsequent nights of sleep (e.g., 48–72 h from learning) seem to maintain the memory and prevent it from decay, with only minimal performance gains, if any at all (Rickard et al., 2008 ; Walker et al., 2003 ). There is no consensus on the optimal delay between training and delayed retention testing, but this length of delay is typically used as it is a more practical way to assess consolidation (Kantak & Winstein, 2012 ). Contextual interference effects are also more apparent after this length of delay (Cahill et al., 2001 ; Perez et al., 2005 ).
Monte Carlo simulation
The implicit-explicit memory distinction may lie on a continuum, with participants having varying amounts of explicit knowledge. However, since we were primarily interested in purely implicit learners, we dichotomized our sample and post hoc sorted participants based on a cutoff determined by a Monte Carlo simulation a priori. This allowed us to determine the number of sequential elements that would be recalled by chance. Subjects would be considered fully implicit learners if their recall was near chance level, and subjects would be considered as having some explicit sequence knowledge if they exceeded this level. This procedure is similar to past research with the SRTT in which a cut-off score based on chance performance was applied to participants’ item recall to define implicit and explicit groups (Robertson et al., 2004 ; Willingham & Goedert-Eschmann, 1999 ). Like the present study, this was to ensure that the individuals in the implicit group had little to no awareness of the sequence.
To determine the amount of explicit knowledge for practiced sequences, we created a score for each participant based on their percentage correct recall for each sequence and averaged the three scores, with 1 as a perfect score. For example, if a subject remembered four sequential items of one sequence (0.5), two sequential items of the second sequence (0.25), and none of the third sequence (0), those three scores would be averaged to create a subject’s “explicit score.” In this example, this subject would have a score of 0.25, meaning that on average, they remembered two items per sequence.
Participants were considered to have only implicit knowledge of practiced sequences if their explicit knowledge of the sequences was near chance levels. To determine chance levels in terms of the number of sequential elements produced, we ran a Monte Carlo simulation. We compared three randomly generated “test” sequences to a set 1,000,000 randomly generated sequences. The test sequences and the comparison set of sequences were in the same format as used in the experiment (i.e., eight item sequences, using each character (U, I, O, P) twice). The percentage similarity score for each test sequence was calculated averaging across all items in the comparison set. We ran this simulation 10,000 times. The generated percentage scores all fell within 0.23–0.26. Thus, on average, we concluded that chance level of performance would be producing about two sequential items per sequence (.25). We considered a participant to have minimal knowledge of the sequences if the participant was able to produce three or fewer sequential items per sequence and used this a-priori cut off to dichotomize our sample. Implicit learners were participants who recalled, on average, 0–3 sequential items per sequence (at chance) while explicit learners recalled 4 or more items per sequence (above chance).
Experiment 1
Participants.
100 right-handed young adults were enrolled in the study (88 female; age 18–48 years, M = 20.9, SD = 4.2). Participants were undergraduate students recruited from UCLA and were given course credit for their participation. All participants gave informed consent using an institutionally approved consent form. Participants were excluded if they had a history of neurological or psychiatric disease or if they were taking neuroactive medication that could affect sensory processing, movement, or cognition. Since the SRTT is a simple task, accuracy likely reflects the degree of participants’ effort, not learning. Thus, participants were excluded if they had an accuracy lower than 80% on either day, in line with previous research (Willingham et al., 2000 ). All participants reported they they were right-handed, although the degree of handedness was not assessed. A total of 17 participants were excluded for either low accuracy ( n = 8), computer error ( n = 5), or failing to complete the experiment ( n = 4). Our final subject pool consisted of 83 right-handed young adults (59 female, age 18–43 years, M = 20.6, SD = 3.2).
Data analysis
Sequence RTs were calculated by summing the eight key press RTs for correct sequences. In line with previous research, only accurate sequences were analyzed (Reber & Squire, 1998 ). We took an average of the last ten sequence RTs per sequence (A, B, and C) for the blocked training condition, for a total of 30 trials. For the interleaved training condition, we took an average of sequence RTs from the last 30 trials. For the blocked testing condition, we used the same procedure but looked at the first ten trials for each sequence, for a total of 30 trials. Similarly, for the interleaved testing condition, we studied the first 30 trials. To measure retention, difference scores were calculated by subtracting the RT of the last 30 trials from Day 1 from the RT of the first 30 trials from Day 2. A negative difference score, reflecting a decrease in RT, indicates improvement (faster performance) from Day 1 to Day 2, while a positive difference score, reflecting an increase in RT, indicates a decline in performance (slower performance) from Day 1 to Day 2. A difference score of zero would indicate successful retention. We assessed learning during the practice phase by looking at sequence RTs over Day 1 (training) in both implicit and explicit learners using the Mann-Kendall test, a nonparametric test for monotonic trends. We also calculated a learning difference score to assess performance at the beginning and end of training. Here, we compared the first 30 trials of interleaved sequences to the last 30 trials, and the first ten of each blocked sequence the last ten of each. Difference scores were calculated similarly to past research (Lin et al., 2010 ; Wymbs & Grafton, 2009 ). We conducted one-sample t-tests for all groups (BI, II, BB, IB ) to assess if difference scores were significantly different from zero, which would indicate an improvement or a decline in performance from Day 1 to Day 2.
To examine whether the different practice conditions yielded different levels of sequence awareness, we compared the average number of sequence items that participants recalled using an independent t-test. We conducted a 2 × 2 × 2 between-subjects ANOVA, after post hoc sorting subjects into implicit and explicit learners based on their recall score. Training condition (Interleaved, Blocked), testing condition (Interleaved, Blocked), and learner type (Implicit/Explicit) were the three factors while the difference score was the dependent measure. In addition, we conducted an ANCOVA with recall score as the covariate before sorting participants into implicit or explicit learners. Like the ANOVA, training and testing condition were the two factors and the difference score was the dependent measure.
On Day 1, a Mann-Whitney test indicated that participants who practiced interleaved sequences were significantly less accurate ( M = 92.34, SD = 4.38) than participants who practiced blocked sequences ( M = 94.22, SD = 4.13; U = 1134.50, p = .013). However, on Day 2, there was no significant difference in accuracy between those who performed interleaved sequences ( M = 94.02, SD = 4.27) and those who performed blocked sequences ( M = 95.59, SD = 2.51; U = 992.50, p = .229). Those who were tested on interleaved sequences either received blocked or interleaved training the day before, however training condition did not impact accuracy on Day 2 ( M II = 93.34, SD II = 4.44; M BI = 94.74, SD BI = 4.06; U = 280.5, p = .234). Similarly, training condition did not impact accuracy on Day 2 for those who were tested on blocked sequences ( M BB = 95.67, SD BB = 2.32; M IB = 95.52, SD IB = 2.72; U = 197.50, p = .968).
Participants who received blocked practice explicitly recalled on average more sequential items per sequence ( M = 4.18, SD = 2.48) than those who had received interleaved practice ( M = 3.17, SD = 2.13); t (81) = -1.996, p = .049). Though 84.3% of participants reported noticing a pattern, only 30.1% of participants correctly noticed there were three sequences. Before categorizing participants into implicit or explicit learners, we conducted a two-way ANCOVA to control for recall score. We found a significant main effect of training condition ( F (1,78) = 38.06, p < .001), a significant main effect of testing condition ( F (1,78) = 10.895, p = .001), and a significant interaction after controlling for recall score ( F (1,78) = 11.565, p = .001). The covariate was not significantly related to performance, indicating that a participants’ knowledge about the sequence had no significant impact on performance or the benefit of interleaved practice ( F (1,78) = 3.02, p = .08). Since our original interest was purely implicit motor sequence learning, we then separated groups based on a cutoff score denoting chance performance. Learners were sorted into two groups based on their explicit recall score: implicit ( n =40) and explicit ( n =43). See Table 1 for breakdown of individual group n s.
For explicit learners, we found significant decreasing monotonic trends for both the interleaved training group ( τ = -.242, p < .0001) as well as the blocked training group ( τ = -.442, p < .0001) (Fig. 2 ). Thus, both groups showed improvement on the SRTT during training on Day 1. This was also true for implicit learners, who also showed significant decreasing monotonic trends for both the interleaved training group ( τ = -.272, p < .0001) as well as the blocked training group ( τ = -.336, p = .001) (Fig. 2 ).

Learning curves during training for Explicit and Implicit Learners, Experiment 1 . Significant decreasing trends in all groups reveal learning over 240 trials
Additionally, we conducted one-sample t-tests to determine if learning difference scores differed significantly from zero, which would indicate a change in performance from the beginning to the end of training on Day 1. Participants in the blocked training condition showed faster RTs at the end of training as compared with the beginning of training ( M = -.9585, SD = .7576); t (39) = -8.00, p <.001, d = -1.27. However, using this measure, there was only a weak trend for a significant difference in RT between the first and last sequences in the interleaved condition ( M = -.199, SD = 0.78). t (42) = -1.6657, p = 0.10, d = -.25, despite the significantly decreasing monotonic trends in RT across the session in the interleaved practice condition for both the implicit and explicit learners. To compare learning difference scores between practice conditions, an independent-samples t-test was conducted. As expected, we found that the blocked training group ( M = -.9585, SD = .7576) did show faster learning during training compared with the interleaved group ( M = -.199, SD = 0.78); t (81) = 4.478, p < .001, d = .98. This is consistent with findings in the CI literature in which blocking stimuli in the absence of high contextual interference facilitates fast performance improvements.
A three-way ANOVA was conducted that examined the effect of training condition, testing condition, and learner type (explicit, implicit) on mean RT difference scores. There was a significant main effect of training condition on difference scores, ( F (1, 75) = 39.539 , p < .001, η 2 = 0.274), with less forgetting from Day 1 to Day 2 for participants who had received interleaved training (Fig. 3 ). Participants who trained in the interleaved condition had a negative difference score, indicating improved performance ( M = -0.313, SD = 0.554). Participants who trained in the blocked condition instead showed a positive difference score, indicating poorer performance from Day 1 to Day 2 ( M = 0.726, SD = 0.938). There was also a significant main effect of testing condition, ( F (1, 75) = 9.538, p = .003, η 2 = 0.066), with greater forgetting for participants who received interleaved testing on Day 2. Participants who received interleaved testing had a mean positive difference score ( M = 0.408, SD = 1.017), while participants who were tested with blocked sequences had a negative mean difference score ( M = -0.049, SD = 0.748).

Mean difference score per condition. Positive difference scores represent increased reaction times (RTs) on Day 2 as compared to Day 1, indicating worse performance. Error bars represent ± SEM
These main effects were qualified by two statistically significant interactions. First, we observed an interaction between the effects of training condition and testing condition on difference scores, F (1,75) = 11.948, p < .001, η 2 = 0.083. An analysis of simple effects showed that testing condition did not significantly affect difference scores when participants were trained in the interleaved condition ( F (1) = 0.027, p = .870). However, testing condition did significantly affect the difference scores when participants were trained in the blocked condition ( F (1) = 20.056, p < .001). Participants who trained with interleaved sequences were able to retain or improve performance regardless of testing condition, while participants who trained with blocked sequences did worse when tested with interleaved sequences.
Additionally, we observed an interaction between the effects of training condition and explicit knowledge on difference scores, F (1, 75) = 4.915, p = .03, η 2 = 0.034. Simple effects analysis showed that learner type did affect difference scores when subjects trained with blocked sequences ( F (1) = 4.529, p = .037), but not when subjects trained with interleaved sequences ( F (1) = 1.082, p = .302). Explicit learners in the blocked training condition had higher difference scores (i.e., more forgetting) than implicit learners, suggesting that explicit learning of the sequences might hinder performance on the delayed test, especially when the sequences were first practiced in a blocked fashion.
In implicit learners, the BI and II groups showed difference scores significantly different than zero. The BI group showed increased RTs on Day 2, indicating forgetting, while the II group showed decreased RTs on Day 2, indicating some consolidation of learning (( t (9) = 3.255, p = .009; t (13) = -3.633, p = .003, respectively). The groups tested with blocked sequences on Day 2 (BB and IB) showed similar RTs across the delay, suggesting good retention of learning across training conditions when sequences were tested in blocks, with no differences between the end of training and the beginning of the test ( t (6) = -.0265, p = .979; t (8) = .119, p = .910, respectively).
In explicit learners, only the BI and IB groups showed difference scores significantly different than zero, in a positive and negative direction, respectively. The BI group showed increased RTs on Day 2 ( t (10) = 5.757, p = .0001), while the IB group demonstrated decreased RTs on Day 2, which reflect faster performance for blocked sequences ( t (11) = -4.849, p = .0005). Difference scores of participants in the II and BB groups did not significantly differ from zero, suggesting good retention from Day 1 ( t (7) = -1.412, p = .201; t (11) = 2.026, p = .068, respectively).
Hours of sleep were assessed; however, participants reported adequate numbers, with little variation ( M = 6.86 , SD = 1.19), and so it was excluded as a covariate.
In this experiment, we used the SRTT to examine the effect of interleaved practice on implicit learning of fine motor sequences after a 1-day delay. We hypothesized that interleaved practice would result in poorer initial performance but superior long-term retention, in line with the CI effect (Shea & Morgan, 1979 ). Consistent with the past findings, we found participants who practiced blocked sequences were much faster than those who practiced interleaved sequences, but they were left unprepared for interleaved sequences at test, especially those with explicit sequence knowledge (Magill & Hall, 1990 ; Shea & Morgan, 1979 ; Wright et al., 2016 ).
We found that interleaved practice may reduce interference from explicit knowledge. An interaction between training condition and learner type (explicit or implicit) revealed that retention was similar for explicit and implicit learners when they practiced interleaved sequences. But for those who practiced blocked sequences, explicit learners were especially impacted, suggesting that explicit knowledge of the sequences may hinder SRTT performance only when sequences are presented in a blocked fashion. This finding is consistent with previous work demonstrating that explicit sequence knowledge can be detrimental to speeded performance in a visuomotor task (Tanaka & Watanabe, 2017 ) as well as on the SRTT (Reber & Squire, 1998 ; but see Willingham et al., 2002 ). It may be that interleaving sequences ameliorates the possible interference that can arise from explicit knowledge, perhaps because practice with high CI encourages general, non-sequence-specific learning that is more immune to intrusions of explicitly learned sequence elements.
The CI effect in motor learning has been reliably demonstrated with explicit sequence learning, but there had been little evidence to suggest that interleaving could benefit sequence learning that occurred without awareness. We found similar results in implicit and explicit learners, in that the CI effect was most pronounced when interleaved sequences were used in the retention test. Even with little to no awareness of any structure within the key presses, those who practiced interleaved sequences showed consolidation of learning when tested with interleaved sequences on Day 2. Those who practiced with blocked sequences were slower when those same sequences were interleaved on Day 2, indicating “forgetting” of sequences that participants were not aware they had learned. These results seem to be in line with the Specificity of Learning hypothesis in that for implicit learners, a consistent practice and testing condition (II) led to better learning than an inconsistent one (BI) (Barnett et al., 1973 ). But those in the IB and BB groups both showed retention, rather than forgetting and consolidation, respectively. This suggests that, similar to other motor studies, our results are more in line with the Variability of Practice hypothesis that emphasizes the role of task variation (e.g., through interleaving) in supporting the learner’s ability to abstract a generalizable schema they can apply to other skills or tasks (Schmidt, 1975 ; Schmidt et al., 1990 ; Shea & Kohl, 1990 ). Blocked practice might result in less flexible learning that is specific to practice conditions, even without conscious awareness of sequences or structure in the task.
In Experiment 1 , interleaved practice led to more flexible retention of practiced sequences in that there was no forgetting for sequences that were presented in either a blocked or interleaved order. In Experiment 2 , we examined whether interleaved practice leads to greater generalization of sequence learning to performance of new sequences. Positive transfer to novel tasks or contexts is a crucial goal in many training situations as one often cannot train on every possible task variation or in every possible context.
Experiment 2
125 right-handed young adults were enrolled in the study (96 female; age 18–30 years, M = 20.6, SD = 1.8). Participants were UCLA undergraduate students and were given course credit for their participation. All participants gave informed consent using an institutionally approved consent form. Participants were excluded if they had a history of neurological or psychiatric disease or if they were taking neuroactive medication that could affect sensory processing, movement, or cognition. All participants reported that they were right-handed, but degree of handedness was not assessed. Participants were excluded if they had an accuracy lower than 80% on either day (Willingham et al., 2000 ). A total of 30 participants were excluded for low accuracy ( n = 16), computer error ( n = 4), and failing to complete the experiment ( n = 10). Our final subject pool consisted of 95 right-handed young adults (72 female, age 18–30 years, M = 20.5, SD = 1.7).
The study design and task is largely the same as Experiment 1 , except that the three sequences presented on Day 2 were novel (e.g., DDD...EEE...FFF for blocked testing or DFEFEDFED….for interleaved testing; Fig. 1 ). Subjects were randomly assigned to one of four training/testing conditions (BB, BI, IB, II). Explicit knowledge of sequences was assessed as in Experiment 1 , except that subjects were asked if they remembered sequences from either Day 1 or Day 2.
Sequence RTs were calculated by summing the eight key press RTs for correct sequences. Data analysis was largely similar to Experiment 1 , except for how the difference score was calculated. To assess transfer, we compared the summed RTs of the first ten presentations of each sequence in the blocked condition or the first 30 sequences in the interleaved condition, and subtracted these from the first ten presentations of each new sequence in the blocked test condition, or the first 30 sequences in the interleaved test condition. Positive transfer would be indicated by faster RTs at the beginning of Day 2 compared to initial performance on Day 1, while negative transfer, or interference, would be indicated by slower RTs. Similar RTs for the beginning of Day 1 and Day 2 would reflect a lack of transfer. We utilized an independent t-test to assess mean items recalled per sequence in both training groups in the explicit knowledge test. We assessed learning during Day 1 training and Day 2 testing using the Mann-Kendall test, a nonparametric test for monotonic trends. Learning difference scores were also calculated by subtracting the RT of the first thirty trials of Day 1 from the RT of the last thirty trials from Day 1.
We also conducted a 2 × 2 between-subjects ANOVA to assess the effect of training and testing conditions on transfer difference scores (beginning of Day 2 – beginning of Day 1). In addition, we conducted an ANCOVA with recall score as the covariate. We also conducted one-sample t-tests to assess which groups (BB, BI, IB, II) had transfer scores that significantly differed from zero.
On Day 1, a Mann-Whitney test indicated that participants who practiced interleaved sequences were significantly less accurate ( M = 91.07, SD = 5.13) than participants who practiced blocked sequences ( M = 94.71, SD = 3.45; U = 1660.00, p < .001). This was also found to be true on Day 2 ( M I = 94.21, SD I = 3.36; M B = 95.72, SD B = 3.07; U = 1455.50, p = .014). Those who were tested on interleaved sequences either received blocked or interleaved training the day before, however training condition did not impact accuracy on Day 2 ( M II = 94.00, SD II = 3.51; M BI = 94.44, SD BI = 3.23; t (48) = 0.47, p = .642). Similarly, training condition did not impact accuracy on Day 2 for those who were tested on blocked sequences ( M BB = 96.30, SD BB = 2.51; M IB = 94.98, SD IB = 3.59; U = 308.50, p = .185).
Explicit recall of sequences was low for both training groups, with no subjects producing more than an average of three sequential items per sequence. On average, subjects recalled fewer than two sequential items per sequence, which is essentially chance recall of sequence elements, indicating that sequence knowledge was substantially implicit. Unlike in Experiment 1 , no participants were excluded from analysis based on substantial explicit sequence knowledge. Though 75.7% of participants reported that they noticed a pattern, only one participant correctly recalled there were six sequences in the entire experiment. There was no significant difference in item recall between participants who trained with interleaved sequences ( M = 1.41 , SD = 1.18) and those who trained with blocked ones ( M = 1.85 , SD = 1.13) ; ( t (96) = -1.8504 , p = .067, d = -0.38). Both groups recalled fewer than two sequential items per sequence on average, demonstrating that all participants learned the sequences implicitly. The reduced amount of explicit sequence knowledge shown by the participants compared to Experiment 1 is likely because Experiment 2 involved six sequences per participant (three sequences on Day 1 followed by three novel sequences on Day 2) and one, not two, sessions of practice on each set of sequences.
We found significant decreasing monotonic trends in RT for both the interleaved training group ( τ = -.431, p < .0001) as well as the blocked training group ( τ = -.39, p < .0001) (Fig. 4 ). This indicates that both groups showed learning of the sequences over Day 1. On Day 2, significant decreasing monotonic trends were found in all groups, except for BI, indicating that blocked training may hinder new learning of interleaved sequences (Table 2 ; Fig. 5 ).

Learning curves during training, Experiment 2 . Significant decreasing trends in both groups reveal learning over 240 trials

Learning curves over Day 2, Experiment 2 . Significant decreasing monotonic trends were found in all groups, except for Blocked-Interleaved, suggesting that blocked training may hinder learning of novel interleaved sequences. Each point is an average of 20 trials
We conducted one-sample t-tests to determine if difference scores from the beginning to the end of training differed significantly from zero, which would indicate a significant change in performance from the beginning to the end of training. Similar to our findings in Experiment 1 , participants in the blocked training condition showed faster RTs at the end of training as compared with the beginning ( M = -1.19; SD = .71); t (48) = -11.74, p < .001; d = -1.68. However, unlike our previous findings, we found that participants in the interleaved training condition did show a significant decrease in RT, demonstrating successful learning as evidenced by a negative difference score ( M = -.31; SD = .64); t (45) = -3.2573, p = .0021, d = -.48. Consistent with our findings in Experiment 1 , we found that the blocked training group ( M = -1.19; SD = .71) showed a greater decrease in RT during training compared to the interleaved group ( M = -.31; SD = .64); t (93) = 6.31, p < .001, d = 1.30.
We next examined transfer learning with a two-way ANOVA of Training condition (Interleaved, Blocked) and Testing condition (Interleaved, Blocked). We found a significant main effect of testing condition, F (1,91) = 9.689, p = .003,η 2 = .092 (Fig. 6 ). Subjects tested with new sequences in the blocked condition showed greater transfer to the new sequences, as evidenced by a negative difference score ( M = -.608, SD = .588). Subjects tested in the interleaved condition also had a negative difference score, but of a smaller magnitude ( M = -.233, SD = .585). There was a trend for a main effect of training condition ( F (1,91) = 3.469, p = .066,η 2 = .033), with participants receiving interleaved training exhibiting numerically greater transfer to new sequences. The interaction between training and testing conditions was not significant ( F (1,91)=1.225, p =.27,η 2 = .012). However, post hoc Tukey tests showed significant differences between BI and IB groups ( M = 0.599, p = .005) as well as between BI and BB groups ( M = 0.509, p = .015). Those in the BI group showed significantly worse transfer performance than those in the IB and BB groups. This indicates that for blocked practice alone, the type of testing condition affects transfer performance. An ANCOVA controlling for recall score reveals a significant main effect of testing condition ( F (1,90) = 8.895, p = .0004,η 2 = .090). Again, the covariate was not found to be significantly related to performance ( F (1,90) = .057, p = .811,η 2 = .0006).

Transfer learning difference score per condition. Note: Negative difference scores indicate faster reaction times (RTs) on Day 2 as compared to Day 1. All groups except for Blocked/Interleaved demonstrated positive transfer to novel sequences. Error bars represent ± SEM
One-sample t-tests revealed that all groups, except for BI, had transfer scores that differed significantly from zero ( M BI = -.054, SD BI =.511, t (23) = -.516 , p =.611). Thus, II, IB, and BB all showed positive transfer to novel sequences ( M II = -.412, SD II = .428, M IB = -.654, SD IB = .919, M BB = -.563, SD BB = .431; p s <.01). In other words, participants in the blocked training condition showed transfer to the new sequences only when they were presented in a blocked order. Participants who received interleaved training showed significantly faster RTs for the new sequences at the beginning of Day 2 compared with the beginning of Day 1 regardless of test condition.
Like the previous experiment, hours of sleep were assessed but were again excluded as a covariate due to sufficient hours of sleep and little variation ( M = 7.07 , SD = 1.49).
In Experiment 2 , we observed implicit sequence learning and transfer for both blocked and interleaved practice conditions in the SRTT. After interleaved practice, there was substantial transfer to performance of new sequences for both testing conditions. After blocked practice, significant positive transfer occurred if new sequences were blocked, but not if they were presented in an interleaved order. This mirrors our results in Experiment 1 in that blocked practice seemed to be vulnerable to testing condition, while interleaved practice prepared participants for both testing conditions. These results suggest that blocked practice of sequences results in implicit learning that is relatively specific to the mode of practice.
While blocked practice did result in positive transfer to new blocked sequences, these participants performed new interleaved sequences at a similar level to their performance at the onset of initial practice. In contrast, interleaved practice of sequences resulted in implicit learning that facilitated performance of new sequences presented in either a blocked or interleaved fashion. In this way, interleaved practice resulted in learning that was general to the SRTT rather than encapsulated in the practiced sequences, similar to previous findings (Müssgens & Ullén, 2015 ). Additionally, prior research using sequential rule paradigms has found that extensive training with a cognitive task was associated with more errors in a transfer task in which the same rules were used but in a different order, suggesting that sequential expectations about a task can interfere with transfer performance (Woltz et al., 2000 ). Blocked practice may thus be more susceptible to violated sequential expectations, resulting in poorer transfer learning, especially when sequences are interleaved.
This result extends recent findings that prior interleaved practice can result in broader learning benefits that are not specific to the practice session. Interleaved practice is usually associated with poorer acquisition and superior retention, however experience with prior interleaved practice may actually improve acquisition of a novel skill (Hodges et al., 2014 ; Kim et al., 2016 , 2018 ). Individuals who underwent prior interleaved practice showed faster acquisition of a novel task as compared with those with prior blocked practice, thereby mitigating the costs normally associated with high CI during learning. This suggests that experience with high CI may facilitate the rate of learning beyond an isolated practice session, perhaps because the learner is able to apply strategies gleaned from interleaved practice to novel motor tasks. It is possible that prior interleaved practice enabled individuals to generate many different motor programs that could aid future learning of similar motor skills (Kim et al., 2018 ). This may also be reflected in our finding that all groups showed learning during Day 2, except those in the BI group. When novel sequences were interleaved at test, prior blocked practice seemed to hinder participants from learning, while those with prior interleaved practice demonstrated successful learning of novel sequences in both testing conditions.
Summary and concluding discussion
We investigated two hypotheses about the effects of interleaved practice on implicit sequence learning. First, we tested whether interleaved practice of sequences leads to greater retention than blocked practice by examining the effect of practice schedule on sequence RT tested the following day after practice. Next, we tested whether interleaved practice of sequences lead to greater positive transfer to novel sequences than blocked practice by examining the effect of practice schedule on performance of novel sequences the day after practice. We found support for the benefit of interleaved practice on both retention and transfer of implicit sequence learning, indicating that the benefit of interleaved practice does not depend on explicit memory retrieval, but also holds for implicit fine motor learning over a delay. Explicit knowledge of the sequence was detrimental to retention when the sequences were blocked, but not when they were interleaved, suggesting that contextual interference may protect against the interference of explicit knowledge on performance.
The SRTT was used here as it is a relatively simple task that has been used extensively to study implicit learning (Robertson, 2007 ). We used eight-item sequences as these are less likely to be learned explicitly than shorter sequences (Meissner et al., 2016 ; Song et al., 2008 ). Nevertheless, in Experiment 1 , where participants practiced the same three sequences for two days, many participants gained at least some awareness of the sequence, particularly in the BB condition. Because more participants became aware of some elements of the sequences in the blocked conditions, it is possible that the unaware participants in the blocked condition differed in some other systematic way from those who gained awareness. It is unclear why some individuals gained partial explicit knowledge of blocked sequences while others did not – perhaps the large number of practice trials (80 per sequence) were repetitive enough for some observant individuals to notice structure in the task (Willingham, 2001 ). The largest group of implicit learners (n=14) was found in the Interleaved/Interleaved condition (Table 1 ). Our results seem consistent with the idea that high CI can encourage implicit learning due to the increased working memory load from frequent task-switching that may make explicit learning more difficult (Rendell et al., 2011 ). In tasks in which explicit knowledge could hinder performance, such as the SRTT (Reber & Squire, 1998 ), CI may facilitate performance by inducing learners to rely on implicit knowledge. Similar results were found by Rendell et al. ( 2011 ), who examined participants’ performance of two gross motor skills of different difficulties while completing a secondary task. Interestingly, participants who practiced a kicking skill with high contextual interference performed exceptionally well under dual-task conditions, suggesting greater implicit learning. The secondary task may require participants to rely on a lesser amount of attentional control and thus these performance gains are due to implicit, rather than explicit, learning. Interleaving tasks may make it more difficult to acquire explicit knowledge, and thus the learner may learn implicitly during acquisition, which in turn maybe more effective for retention and transfer performance. Notably, this result only applied to the more complex skill of kicking, so task difficulty is an important consideration when examining CI (Albaret & Thon, 1998 ; Farrow & Buszard, 2017 ; Guadagnoli & Lee, 2004 ). The Challenge Point Framework predicts that the strength of the CI effect is partially determined by task difficulty; namely that it is more robust with low difficulty tasks (Guadagnoli & Lee, 2004 ). Since Rendell et al. ( 2011 ) only observed the CI effect in the more challenging task, these results seem inconsistent with this framework. In the present study, we used a low difficulty task and were able to observe benefits of CI during practice. Future research should aim to clarify these disparate findings and specifically manipulate task difficulty under varying levels of contextual interference. It is possible that with more complex sequences there would be little or no benefit of CI.
Recent research indicates that frequent error processing in addition to task switching increases cognitive effort and may encourage implicit learning (Broadbent et al., 2017 ). In both experiments presented here we found that those in the interleaved condition were less accurate than those in the blocked condition on Day 1, which may lend some credence to the theory that frequent error processing may occur with high CI (Broadbent et al., 2017 ). However, on Day 2 of Experiment 1 , accuracy did not differ between the two conditions, and we did not observe the costs normally associated with interleaved practice, consistent with past research (Hodges et al., 2014 ) which may reflect a general learning benefit of interleaving that results in improved skill acquisition (Kim et al., 2016 , 2018 ).
In Experiment 2 , we focused on transfer to new sequences. This type of transfer may be conceptually related to playing a new piece of music after extensive practice of a different piece. Implicit learning has been thought to be inflexible and not amenable to transfer (Dienes & Berry, 1997 ; Sanchez et al., 2015 ), though our results suggest that positive transfer of implicit motor sequences may be facilitated by introducing high CI during acquisition. The present results are the first to show a benefit for interleaving in the positive transfer of learned sequences in which the lack of awareness of learning was assessed, indicating that explicit processes may not be required to observe this effect. Past research has found that explicit knowledge can reduce errors but increase RTs, hindering transfer performance overall (Benson et al., 2011 ; Tanaka & Watanabe, 2017 ). However, error processing may be an important component of the learning process as errors allow learners to compare between the actual versus expected outcomes of response selection, which can then inform hypotheses and rules that learners may generate to improve performance (Maxwell et al., 2001 ; Rabbitt, 1966 ). Thus, fewer errors due to explicit knowledge may actually hinder successful long term retention despite superior acquisition performance. As discussed above, this is in line with the idea that interleaving increases working memory load, presumably by task switching and error processing, which encourages an implicit form of learning (Broadbent et al., 2017 ; Rendell et al., 2011 ).
Though promising, our results should be cautiously interpreted due to some limitations. Sample size for both experiments was analogous to similar studies, but the substantial number of explicit learners in Experiment 1 led to a smaller sample size for the implicit learner group. However, we felt that dichotomizing was essential as we were interested in whether the CI effect persists in the absence of explicit knowledge, as implicit learning is an important component of long-term skill retention in both healthy adults (Howard et al., 2004 ; Mazzoni & Krakauer, 2006 ) and for clinical populations with explicit memory deficits (Curran, 1997 ; Gabrieli et al., 1993 ) or those with chronic stroke (Boyd & Winstein, 2004 ; Dimyan & Cohen, 2011 ; Wadden et al., 2017 ). Though prior literature has used similar methods to exclude those with explicit awareness, we acknowledge that this categorization does not address the question of how the effects of interleaving are impacted by increasing levels of awareness of learning. Additionally, some participants who were classified as explicit learners may have only gained explicit knowledge of the sequences towards the end of the retention test, and thus may have learned the sequences implicitly during Day 1. Future research could examine the effects of CI when learning is more clearly implicit in all participants, such as learning with a concurrent task (Grafton et al., 1995 ; Nejati et al., 2008 ) or a more complex probabilistic sequence (Du et al., 2016 ; Song et al., 2007 ).
Fatigue was a potential factor that impacted performance given the large number of trials during training and testing. Extended practice may cause fatigue and participants might fail to show learning despite successful skill acquisition (Eysenck, 1965 ). Performance might improve if fatigue could be mitigated with intervals of rest, but previous work demonstrated that adding breaks to the SRTT did little to change performance in implicit learners (Robertson et al., 2004 ). Additional evidence suggests measuring motor skill learning via RT difference scores is unaffected by fatigue (Heuer et al., 1998 ). However, it is possible that fatigue impacted participants’ motivation or effort, given the number of participants who were excluded based on low accuracy or failing to complete the experiment, despite the SRTT’s relative simplicity. Future research may utilize fewer trials, as implicit sequence transfer may occur even with a shorter learning period (Tanaka & Watanabe, 2015 ).
In sum, the present results add to the literature showing the benefits of interleaved practice on learning and transfer and extend them by providing evidence that this effect is also observed in implicit fine motor sequence learning. Blocked practice of sequences in real world skills, such as repetitively playing single pieces when learning an instrument, may appear to be effective in that performance will improve within the practice session. However, this type of practice may not be as effective for mastering the instrument as interleaved practice of different pieces. Furthermore, these results suggest that patients with deficits in explicit memory can still benefit from high CI to successfully learn and retain novel motor skills.
Open practices statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. Neither of the experiments were preregistered.
Abrahamse, E., Ruitenberg, M., De Kleine, E., & Verwey, W. B. (2013). Control of automated behavior: Insights from the discrete sequence production task. Frontiers in Human Neuroscience , 7 . https://doi.org/10.3389/fnhum.2013.00082
Albaret, J.-M., & Thon, B. (1998). Differential effects of task complexity on contextual interference in a drawing task. Acta Psychologica , 100 (1), 9–24. https://doi.org/10.1016/S0001-6918(98)00022-5
Article PubMed Google Scholar
Bangert, M., Wiedemann, A., & Jabusch, H.-C. (2014). Effects of variability of practice in music: A pilot study on fast goal-directed movements in pianists. Frontiers in Human Neuroscience , 8 . https://doi.org/10.3389/fnhum.2014.00598
Barnett, M. L., Ross, D., Schmidt, R. A., & Todd, B. (1973). Motor Skills Learning and the Specificity of Training Principle. Research Quarterly. American Association for Health, Physical Education and Recreation , 44 (4), 440–447. https://doi.org/10.1080/10671188.1973.10615224
Article Google Scholar
Battig, W. (1966). Facilitation and Interference. In E. A. Bilodeau (Ed.), Acquisition of Skill (pp. 215–244). Academic Press.
Benson, B. L., Anguera, J. A., & Seidler, R. D. (2011). A spatial explicit strategy reduces error but interferes with sensorimotor adaptation. Journal of Neurophysiology , 105 (6), 2843–2851. https://doi.org/10.1152/jn.00002.2011
Article PubMed PubMed Central Google Scholar
Bjork, R. A. (1994). Memory and metamemory considerations in the training of human beings. In Metacognition: Knowing about knowing (pp. 185–205). The MIT Press.
Bjork, R. A., & Kroll, J. F. (2015). Desirable Difficulties in Vocabulary Learning. The American Journal of Psychology , 128 (2), 241–252.
Bo, J., & Seidler, R. D. (2009). Visuospatial Working Memory Capacity Predicts the Organization of Acquired Explicit Motor Sequences. Journal of Neurophysiology , 101 (6), 3116–3125. https://doi.org/10.1152/jn.00006.2009
Bönstrup, M., Iturrate, I., Thompson, R., Cruciani, G., Censor, N., & Cohen, L. G. (2019). A Rapid Form of Offline Consolidation in Skill Learning. Current Biology , 29 (8), 1346-1351.e4. https://doi.org/10.1016/j.cub.2019.02.049
Boyd, L. A., & Winstein, C. J. (2004). Providing explicit information disrupts implicit motor learning after basal ganglia stroke. Learning & Memory (Cold Spring Harbor, N.Y.) , 11 (4), 388–396. https://doi.org/10.1101/lm.80104
Brady, F. (2004). Contextual interference: A meta-analytic study. Perceptual and Motor Skills , 99 (1), 116–126. https://doi.org/10.2466/pms.99.1.116-126
Broadbent, D. P., Causer, J., Williams, A. M., & Ford, P. R. (2017). The role of error processing in the contextual interference effect during the training of perceptual-cognitive skills. Journal of Experimental Psychology: Human Perception and Performance , 43 (7), 1329. https://doi.org/10.1037/xhp0000375
Cahill, L., McGaugh, J. L., & Weinberger, N. M. (2001). The neurobiology of learning and memory: Some reminders to remember. Trends in Neurosciences , 24 (10), 578–581. https://doi.org/10.1016/S0166-2236(00)01885-3
Curran, T. (1997). Higher-Order Associative Learning in Amnesia: Evidence from the Serial Reaction Time Task. Journal of Cognitive Neuroscience , 9 (4), 522–533. https://doi.org/10.1162/jocn.1997.9.4.522
Dang, K. V., Parvin, D. E., & Ivry, R. B. (2019). Exploring Contextual Interference in Implicit and Explicit Motor Learning. BioRxiv , 644211. https://doi.org/10.1101/644211
Dienes, Z., & Berry, D. (1997). Implicit learning: Below the subjective threshold. Psychonomic Bulletin & Review , 4 (1), 3–23. https://doi.org/10.3758/BF03210769
Dimyan, M. A., & Cohen, L. G. (2011). Neuroplasticity in the context of motor rehabilitation after stroke. Nature Reviews. Neurology , 7 (2), 76–85. https://doi.org/10.1038/nrneurol.2010.200
Du, Y., Prashad, S., Schoenbrun, I., & Clark, J. E. (2016). Probabilistic Motor Sequence Yields Greater Offline and Less Online Learning than Fixed Sequence. Frontiers in Human Neuroscience , 10 . https://doi.org/10.3389/fnhum.2016.00087
Eysenck, H. J. (1965). A three-factor theory of reminiscence. British Journal of Psychology , 56 (2–3), 163–182. https://doi.org/10.1111/j.2044-8295.1965.tb00956.x
Farrow, D., & Buszard, T. (2017). Chapter 5—Exploring the applicability of the contextual interference effect in sports practice. In M. R. Wilson, V. Walsh, & B. Parkin (Eds.), Progress in Brain Research (Vol. 234, pp. 69–83). Elsevier. https://doi.org/10.1016/bs.pbr.2017.07.002
French, K. E., Rink, J. E., & Werner, P. H. (1990). Effects of contextual interference on retention of three volleyball skills. Perceptual and Motor Skills , 71 (1), 179–186. https://doi.org/10.2466/PMS.71.4.179-186
Frensch, P. A., & Rünger, D. (2003). Implicit Learning: Current Directions in Psychological Science . https://journals.sagepub.com/doi/10.1111/1467-8721.01213
Gabrieli, J. D., Corkin, S., Mickel, S. F., & Growdon, J. H. (1993). Intact acquisition and long-term retention of mirror-tracing skill in Alzheimer’s disease and in global amnesia. Behavioral Neuroscience , 107 (6), 899–910.
Goode, S., & Magill, R. A. (1986). Contextual Interference Effects in Learning Three Badminton Serves. Research Quarterly for Exercise and Sport , 57 (4), 308–314. https://doi.org/10.1080/02701367.1986.10608091
Grafton, S. T., Hazeltine, E., & Ivry, R. (1995). Functional Mapping of Sequence Learning in Normal Humans. Journal of Cognitive Neuroscience , 7 (4), 497–510. https://doi.org/10.1162/jocn.1995.7.4.497
Guadagnoli, M. A., & Lee, T. D. (2004). Challenge point: A framework for conceptualizing the effects of various practice conditions in motor learning. Journal of Motor Behavior , 36 (2), 212–224. https://doi.org/10.3200/JMBR.36.2.212-224
Heuer, H., Spijkers, W., Kiesswetter, E., & Schmidtke, V. (1998). Effects of sleep loss, time of day, and extended mental work on implicit and explicit learning of sequences. Journal of Experimental Psychology: Applied , 4 (2), 139–162. https://doi.org/10.1037/1076-898X.4.2.139
Hodges, N. J., Lohse, K. R., Wilson, A., Lim, S. B., & Mulligan, D. (2014). Exploring the Dynamic Nature of Contextual Interference: Previous Experience Affects Current Practice But Not Learning. Journal of Motor Behavior, 46 (6), 455–467.
Howard, D. V., Howard, J. H., Japikse, K., DiYanni, C., Thompson, A., & Somberg, R. (2004). Implicit sequence learning: Effects of level of structure, adult age, and extended practice. Psychology and Aging , 19 (1), 79–92. https://doi.org/10.1037/0882-7974.19.1.79
Kang, S. H. K., & Pashler, H. (2012). Learning Painting Styles: Spacing is Advantageous when it Promotes Discriminative Contrast: Spacing promotes contrast. Applied Cognitive Psychology , 26 (1), 97–103. https://doi.org/10.1002/acp.1801
Kantak, S. S., & Winstein, C. J. (2012). Learning-performance distinction and memory processes for motor skills: A focused review and perspective. Behavioural Brain Research , 228 (1), 219–231. https://doi.org/10.1016/j.bbr.2011.11.028
Keele, S. W., Ivry, R., Mayr, U., Hazeltine, E., & Heuer, H. (2003). The cognitive and neural architecture of sequence representation. Psychological Review , 110 (2), 316–339. https://doi.org/10.1037/0033-295X.110.2.316
Kelso, J. A. S., & Zanone, P.-G. (2002). Coordination dynamics of learning and transfer across different effector systems. Journal of Experimental Psychology: Human Perception and Performance , 28 (4), 776–797. https://doi.org/10.1037/0096-1523.28.4.776
Kim, T., Chen, J., Verwey, W. B., & Wright, D. L. (2018). Improving novel motor learning through prior high contextual interference training. Acta Psychologica , 182 , 55–64. https://doi.org/10.1016/j.actpsy.2017.11.005
Kim, T., Rhee, J., & Wright, D. L. (2016). Allowing time to consolidate knowledge gained through random practice facilitates later novel motor sequence acquisition. Acta Psychologica , 163 , 153–166. https://doi.org/10.1016/j.actpsy.2015.11.012
Kim, T, & Wright, D. L. (2020). Transcranial Direct Current Stimulation of Supplementary Motor Region Impacts the Effectiveness of Interleaved and Repetitive Practice Schedules for Retention of Motor Skills. Neuroscience , 435 , 58–72. https://doi.org/10.1016/j.neuroscience.2020.03.043
Lee, T. D., Magill, R. A., & Weeks, D. J. (1985). Influence of practice schedule on testing schema theory predictions in adults. Journal of Motor Behavior , 17 (3), 283–299. https://doi.org/10.1080/00222895.1985.10735350
Lee, T., & Simon, D. (2004). Contextual interference. In A. Williams & N. Hodges (Eds.), Skill acquisition in sport: Research, theory and practice (pp. 29–44). Routledge.
Lee, Timothy D., & Magill, R. A. (1983). The locus of contextual interference in motor-skill acquisition. Journal of Experimental Psychology: Learning, Memory, and Cognition , 9 (4), 730–746. https://doi.org/10.1037/0278-7393.9.4.730
Li, Y., & Wright, D. L. (2000). An Assessment of the Attention Demands during Random- and Blocked-Practice Schedules. The Quarterly Journal of Experimental Psychology Section A , 53 (2), 591–606. https://doi.org/10.1080/713755890
Lin, C. H., Fisher, B. E., Winstein, C. J., Wu, A. D., & Gordon, J. (2008). Contextual Interference Effect: Elaborative Processing or Forgetting—Reconstruction? A Post Hoc Analysis of Transcranial Magnetic Stimulation—Induced Effects on Motor Learning. Journal of Motor Behavior , 40 (6), 578–586. https://doi.org/10.3200/JMBR.40.6.578-586
Lin, C.-H. J., Knowlton, B. J., Chiang, M.-C., Iacoboni, M., Udompholkul, P., & Wu, A. D. (2011). Brain-behavior correlates of optimizing learning through interleaved practice. NeuroImage , 56 (3), 1758–1772. https://doi.org/10.1016/j.neuroimage.2011.02.066
Lin, C.-H. J., Wu, A. D., Udompholkul, P., & Knowlton, B. J. (2010). Contextual interference effects in sequence learning for young and older adults. Psychology and Aging , 25 (4), 929–939. https://doi.org/10.1037/a0020196
Magill, R. A., & Hall, K. G. (1990). A review of the contextual interference effect in motor skill acquisition. Human Movement Science , 9 (3), 241–289. https://doi.org/10.1016/0167-9457(90)90005-X
Maxwell, J. P., Masters, R. S., Kerr, E., & Weedon, E. (2001). The implicit benefit of learning without errors. The Quarterly Journal of Experimental Psychology. A, Human Experimental Psychology , 54 (4), 1049–1068. https://doi.org/10.1080/713756014
Mazzoni, P., & Krakauer, J. W. (2006). An Implicit Plan Overrides an Explicit Strategy during Visuomotor Adaptation. The Journal of Neuroscience , 26 (14), 3642–3645. https://doi.org/10.1523/JNEUROSCI.5317-05.2006
Meira, C. M., Jr., & Tani, G. (2001). The contextual interference effect in acquisition of dart-throwing skill tested on a transfer test with extended trials. Perceptual and Motor Skills , 92 (3,Pt1), 910–918. https://doi.org/10.2466/PMS.92.3.910-918
Meissner, S. N., Keitel, A., Südmeyer, M., & Pollok, B. (2016). Implicit Motor Sequence Learning and Working Memory Performance Changes Across the Adult Life Span. Frontiers in Aging Neuroscience , 8 . https://doi.org/10.3389/fnagi.2016.00089
Menayo, R., Moreno, F. J., Sabido, R., Fuentes, J. P., & García, J. A. (2010). Simultaneous treatment effects in learning four tennis shots in contextual interference conditions. Perceptual and Motor Skills , 110 (2), 661–673. https://doi.org/10.2466/PMS.110.2.661-673
Morris, C. D., Bransford, J. D., & Franks, J. J. (1977). Levels of processing versus transfer appropriate processing. Journal of Verbal Learning & Verbal Behavior , 16 (5), 519–533. https://doi.org/10.1016/S0022-5371(77)80016-9
Müssgens, D. M., & Ullén, F. (2015). Transfer in Motor Sequence Learning: Effects of Practice Schedule and Sequence Context. Frontiers in Human Neuroscience , 9 . https://doi.org/10.3389/fnhum.2015.00642
Nader, K., Schafe, G. E., & Le Doux, J. E. (2000). Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature , 406 (6797), 722–726. https://doi.org/10.1038/35021052
Nejati, V., Farshi, M. T. G., Ashayeri, H., & Aghdasi, M. T. (2008). Dual task interference in implicit sequence learning by young and old adults. International Journal of Geriatric Psychiatry , 23 (8), 801–804. https://doi.org/10.1002/gps.1976
Newell, K. (1996). Change in Movement and Skill: Learning, Retention, and Transfer. In M. Latash & M. Turvey (Eds.), Dexterity and Its Development .
Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learning: Evidence from performance measures. Cognitive Psychology , 19 (1), 1–32. https://doi.org/10.1016/0010-0285(87)90002-8
Obayashi, S. (2004). Possible mechanism for transfer of motor skill learning: Implication of the cerebellum. The Cerebellum , 3 (4), 204–211. https://doi.org/10.1080/14734220410018977
Perez, C. R., Meira, C. M., & Tani, G. (2005). Does the Contextual Interference Effect Last over Extended Transfer Trials? Perceptual and Motor Skills , 100 (1), 58–60. https://doi.org/10.2466/pms.100.1.58-60
Rabbitt, P. M. (1966). Errors and error correction in choice-response tasks. Journal of Experimental Psychology , 71 (2), 264–272. https://doi.org/10.1037/h0022853
Reber, P. J., & Squire, L. R. (1994). Parallel brain systems for learning with and without awareness. Learning & Memory (Cold Spring Harbor, N.Y.) , 1 (4), 217–229.
Google Scholar
Reber, P. J., & Squire, L. R. (1998). Encapsulation of implicit and explicit memory in sequence learning. Journal of Cognitive Neuroscience , 10 (2), 248–263. https://doi.org/10.1162/089892998562681
Rendell, M. A., Masters, R. S. W., Farrow, D., & Morris, T. (2011). An implicit basis for the retention benefits of random practice. Journal of Motor Behavior , 43 (1), 1–13. https://doi.org/10.1080/00222895.2010.530304
Rickard, T. C., Cai, D. J., Rieth, C. A., Jones, J., & Ard, M. C. (2008). Sleep does not enhance motor sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition , 34 (4), 834–842. https://doi.org/10.1037/0278-7393.34.4.834
Robertson, E. M. (2007). The serial reaction time task: Implicit motor skill learning? The Journal of Neuroscience: The Official Journal of the Society for Neuroscience , 27 (38), 10073–10075. https://doi.org/10.1523/JNEUROSCI.2747-07.2007
Robertson, E. M., Pascual-Leone, A., & Press, D. Z. (2004). Awareness modifies the skill-learning benefits of sleep. Current Biology: CB , 14 (3), 208–212. https://doi.org/10.1016/j.cub.2004.01.027
Russell, D. M., & Newell, K. M. (2007). How persistent and general is the contextual interference effect? Research Quarterly for Exercise and Sport , 78 (4), 318–327. https://doi.org/10.1080/02701367.2007.10599429
Sanchez, D. J., Yarnik, E. N., & Reber, P. J. (2015). Quantifying transfer after perceptual-motor sequence learning: How inflexible is implicit learning? Psychological Research , 79 (2), 327–343. https://doi.org/10.1007/s00426-014-0561-9
Schmidt, R. A. (1975). A schema theory of discrete motor skill learning. Psychological Review , 82 (4), 225–260. https://doi.org/10.1037/h0076770
Schmidt, R. A., Lange, C., & Young, D. E. (1990). Optimizing summary knowledge of results for skill learning. Human Movement Science , 9 (3), 325–348. https://doi.org/10.1016/0167-9457(90)90007-Z
Schmidt, R. A., & Lee, T. D. (2005). Motor control and learning: A behavioral emphasis, 4th ed . Human Kinetics.
Sekiya, H. (2006). Contextual interference in implicit and explicit motor learning. Perceptual and Motor Skills , 103 (2), 333–343. https://doi.org/10.2466/pms.103.2.333-343
Shanks, D. R., & St. John, M. F. (1994). Characteristics of dissociable human learning systems. Behavioral and Brain Sciences , 17 (3), 367–395. https://doi.org/10.1017/S0140525X00035032
Shea, C. H., & Kohl, R. M. (1990). Specificity and variability of practice. Research Quarterly for Exercise and Sport , 61 (2), 169–177. https://doi.org/10.1080/02701367.1990.10608671
Shea, J. B., Hunt, J. P., & Zimny, S. T. (1985). Representational Structure and Strategic Processes for Movement Production. In D. Goodman, R. B. Wilberg, & I. M. Franks (Eds.), Advances in Psychology (Vol. 27, pp. 55–87). North-Holland. https://doi.org/10.1016/S0166-4115(08)62530-3
Shea, J. B., & Zimny, S. T. (1983). Context Effects in Memory and Learning Movement Information. In R. A. Magill (Ed.), Advances in Psychology (Vol. 12, pp. 345–366). North-Holland. https://doi.org/10.1016/S0166-4115(08)61998-6
Shea, J. B., & Morgan, R. (1979). Contextual Interference Effects on the Acquisition, Retention, and Transfer of a Motor Skill. Journal of Experimental Psychology: Human Learning and Memory , 5 (2), 179–187.
Shimizu, R. E., Wu, A. D., & Knowlton, B. J. (2016). Cerebellar activation during motor sequence learning is associated with subsequent transfer to new sequences. Behavioral Neuroscience , 130 (6), 572–584. https://doi.org/10.1037/bne0000164
Shmuelof, L., & Krakauer, J. W. (2014). Recent insights into perceptual and motor skill learning. Frontiers in Human Neuroscience, 8 . https://doi.org/10.3389/fnhum.2014.00683
Soderstrom, N. C., & Bjork, R. A. (2015). Learning Versus Performance: An Integrative Review. Perspectives on Psychological Science , 10 (2), 176–199. https://doi.org/10.1177/1745691615569000
Song, S., Howard, J. H., & Howard, D. V. (2007). Implicit probabilistic sequence learning is independent of explicit awareness. Learning & Memory , 14 (3), 167–176. https://doi.org/10.1101/lm.437407
Song, S., Howard, J. H., & Howard, D. V. (2008). Perceptual sequence learning in a serial reaction time task. Experimental Brain Research. Experimentelle Hirnforschung. Experimentation Cerebrale , 189 (2), 145–158. https://doi.org/10.1007/s00221-008-1411-z
Squire, L. R., Genzel, L., Wixted, J. T., & Morris, R. G. (2015). Memory consolidation. Cold Spring Harbor Perspectives in Biology , 7 (8), a021766. https://doi.org/10.1101/cshperspect.a021766
Stark-Inbar, A., Raza, M., Taylor, J. A., & Ivry, R. B. (2017). Individual differences in implicit motor learning: Task specificity in sensorimotor adaptation and sequence learning. Journal of Neurophysiology , 117 (1), 412–428. https://doi.org/10.1152/jn.01141.2015
Tanaka, K., & Watanabe, K. (2015). Effects of learning duration on implicit transfer. Experimental Brain Research , 233 (10), 2767–2776. https://doi.org/10.1007/s00221-015-4348-z
Tanaka, K., & Watanabe, K. (2017). Explicit instruction of rules interferes with visuomotor skill transfer. Experimental Brain Research , 235 (6), 1689–1700. https://doi.org/10.1007/s00221-017-4933-4
Travlos, A. K. (2010). Specificity and Variability of Practice, and Contextual Interference in Acquisition and Transfer of an Underhand Volleyball Serve. Perceptual and Motor Skills , 110 (1), 298–312. https://doi.org/10.2466/pms.110.1.298-312
Wadden, K. P., De Asis, K., Mang, C. S., Neva, J. L., Peters, S., Lakhani, B., & Boyd, L. A. (2017). Predicting Motor Sequence Learning in Individuals with Chronic Stroke. Neurorehabilitation and Neural Repair , 31 (1), 95–104. https://doi.org/10.1177/1545968316662526
Walker, M. P., Brakefield, T., Allan Hobson, J., & Stickgold, R. (2003). Dissociable stages of human memory consolidation and reconsolidation. Nature , 425 (6958), 616–620. https://doi.org/10.1038/nature01930
Walker, M. P., Brakefield, T., Morgan, A., Hobson, J. A., & Stickgold, R. (2002). Practice with sleep makes perfect: Sleep-dependent motor skill learning. Neuron , 35 (1), 205–211.
Willingham, D. B. (2001). Becoming aware of motor skill. Trends in Cognitive Sciences , 5 (5), 181–182. https://doi.org/10.1016/S1364-6613(00)01652-1
Willingham, D. B., & Goedert-Eschmann, K. (1999). The Relation Between Implicit and Explicit Learning: Evidence for Parallel Development. Psychological Science , 10 (6), 531–534. https://doi.org/10.1111/1467-9280.00201
Willingham, D. B., Salidis, J., & Gabrieli, J. D. E. (2002). Direct Comparison of Neural Systems Mediating Conscious and Unconscious Skill Learning. Journal of Neurophysiology , 88 (3), 1451–1460. https://doi.org/10.1152/jn.2002.88.3.1451
Willingham, D. B., Wells, L. A., Farrell, J. M., & Stemwedel, M. E. (2000). Implicit motor sequence learning is represented in response locations. Memory & Cognition , 28 (3), 366–375. https://doi.org/10.3758/BF03198552
Woltz, D. J., Gardner, M. K., & Bell, B. G. (2000). Negative transfer errors in sequential cognitive skills: Strong-but-wrong sequence application. Journal of Experimental Psychology: Learning, Memory, and Cognition , 26 (3), 601–625. https://doi.org/10.1037/0278-7393.26.3.601
Wright, D., Verwey, W., Buchanen, J., Chen, J., Rhee, J., & Immink, M. (2016). Consolidating behavioral and neurophysiologic findings to explain the influence of contextual interference during motor sequence learning. Psychonomic Bulletin & Review , 23 (1), 1–21. https://doi.org/10.3758/s13423-015-0887-3
Wymbs, N. F., & Grafton, S. T. (2009). Neural Substrates of Practice Structure That Support Future Off-Line Learning. Journal of Neurophysiology , 102 (4), 2462–2476. https://doi.org/10.1152/jn.00315.2009
Download references
Author information
Authors and affiliations.
Department of Psychology, University of California, Los Angeles, Los Angeles, CA, 90095, USA
Julia M. Schorn & Barbara J. Knowlton
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Julia M. Schorn .
Ethics declarations
Conflict of interest.
We have no known conflict of interest to disclose.
Author note
This research was supported in part by the National Science Foundation (BCS- 1634157 to B.K.) and UCLA’s Graduate Research Mentorship Fellowship (to J.S.).
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Schorn, J.M., Knowlton, B.J. Interleaved practice benefits implicit sequence learning and transfer. Mem Cogn 49 , 1436–1452 (2021). https://doi.org/10.3758/s13421-021-01168-z
Download citation
Accepted : 09 March 2021
Published : 01 April 2021
Issue Date : October 2021
DOI : https://doi.org/10.3758/s13421-021-01168-z
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Contextual interference
- Serial reaction time task
- Implicit learning
- Find a journal
- Publish with us
- Track your research
- More from M-W
- To save this word, you'll need to log in. Log In
Definition of hypothesis
Did you know.
The Difference Between Hypothesis and Theory
A hypothesis is an assumption, an idea that is proposed for the sake of argument so that it can be tested to see if it might be true.
In the scientific method, the hypothesis is constructed before any applicable research has been done, apart from a basic background review. You ask a question, read up on what has been studied before, and then form a hypothesis.
A hypothesis is usually tentative; it's an assumption or suggestion made strictly for the objective of being tested.
A theory , in contrast, is a principle that has been formed as an attempt to explain things that have already been substantiated by data. It is used in the names of a number of principles accepted in the scientific community, such as the Big Bang Theory . Because of the rigors of experimentation and control, it is understood to be more likely to be true than a hypothesis is.
In non-scientific use, however, hypothesis and theory are often used interchangeably to mean simply an idea, speculation, or hunch, with theory being the more common choice.
Since this casual use does away with the distinctions upheld by the scientific community, hypothesis and theory are prone to being wrongly interpreted even when they are encountered in scientific contexts—or at least, contexts that allude to scientific study without making the critical distinction that scientists employ when weighing hypotheses and theories.
The most common occurrence is when theory is interpreted—and sometimes even gleefully seized upon—to mean something having less truth value than other scientific principles. (The word law applies to principles so firmly established that they are almost never questioned, such as the law of gravity.)
This mistake is one of projection: since we use theory in general to mean something lightly speculated, then it's implied that scientists must be talking about the same level of uncertainty when they use theory to refer to their well-tested and reasoned principles.
The distinction has come to the forefront particularly on occasions when the content of science curricula in schools has been challenged—notably, when a school board in Georgia put stickers on textbooks stating that evolution was "a theory, not a fact, regarding the origin of living things." As Kenneth R. Miller, a cell biologist at Brown University, has said , a theory "doesn’t mean a hunch or a guess. A theory is a system of explanations that ties together a whole bunch of facts. It not only explains those facts, but predicts what you ought to find from other observations and experiments.”
While theories are never completely infallible, they form the basis of scientific reasoning because, as Miller said "to the best of our ability, we’ve tested them, and they’ve held up."
- proposition
- supposition
hypothesis , theory , law mean a formula derived by inference from scientific data that explains a principle operating in nature.
hypothesis implies insufficient evidence to provide more than a tentative explanation.
theory implies a greater range of evidence and greater likelihood of truth.
law implies a statement of order and relation in nature that has been found to be invariable under the same conditions.
Examples of hypothesis in a Sentence
These examples are programmatically compiled from various online sources to illustrate current usage of the word 'hypothesis.' Any opinions expressed in the examples do not represent those of Merriam-Webster or its editors. Send us feedback about these examples.
Word History
Greek, from hypotithenai to put under, suppose, from hypo- + tithenai to put — more at do
1641, in the meaning defined at sense 1a
Phrases Containing hypothesis
- counter - hypothesis
- nebular hypothesis
- null hypothesis
- planetesimal hypothesis
- Whorfian hypothesis
Articles Related to hypothesis

This is the Difference Between a...
This is the Difference Between a Hypothesis and a Theory
In scientific reasoning, they're two completely different things
Dictionary Entries Near hypothesis
hypothermia
hypothesize
Cite this Entry
“Hypothesis.” Merriam-Webster.com Dictionary , Merriam-Webster, https://www.merriam-webster.com/dictionary/hypothesis. Accessed 22 Jun. 2024.
Kids Definition
Kids definition of hypothesis, medical definition, medical definition of hypothesis, more from merriam-webster on hypothesis.
Nglish: Translation of hypothesis for Spanish Speakers
Britannica English: Translation of hypothesis for Arabic Speakers
Britannica.com: Encyclopedia article about hypothesis
Subscribe to America's largest dictionary and get thousands more definitions and advanced search—ad free!

Can you solve 4 words at once?
Word of the day.
See Definitions and Examples »
Get Word of the Day daily email!
Popular in Grammar & Usage
Plural and possessive names: a guide, more commonly misspelled words, your vs. you're: how to use them correctly, every letter is silent, sometimes: a-z list of examples, more commonly mispronounced words, popular in wordplay, 8 words for lesser-known musical instruments, birds say the darndest things, 10 words from taylor swift songs (merriam's version), 10 scrabble words without any vowels, 12 more bird names that sound like insults (and sometimes are), games & quizzes.

IMAGES
VIDEO
COMMENTS
Two theories have been proposed to explain the underlying mechanisms of the CI effect, namely the elaborative processing hypothesis and the action plan reconstruction hypothesis.Both theories detail how greater cognitive effort occurs during random compared with blocked ordered practice due to task switching (Lee, 2012).Cognitive effort is the mental work involved in selecting and executing ...
With regard to the previously discussed behaviorally based theoretical accounts of the CI effect, i.e., the elaboration and action-plan reconstruction hypotheses, the following is noteworthy: even though our imaging findings do not provide unequivocal evidence for one of both hypotheses, they seem to favor the elaboration hypothesis (Shea and ...
The reconstruction hypothesis (Lee and Magill, 1983, Lee and Magill, 1985) relies on verbal learning research that addresses the spacing effect ... The basic idea is that random practice causes the learner to forget some task-specific information of the action plan previously encoded, and consequently, forces the learner to engage in a ...
However, Lee and Magill proposed the action-plan reconstruction hypothesis stating that action plans will be forgotten because of alternating trials following a random practice schedule. Thus, high amounts of CI will result in more effortful reconstructive processing whereas the action plan in blocked practice will be remembered.
2)The Action Plan Reconstruction Hypothesis (Lee & Magill, 1985). In the Action Plan Reconstruction Hypothesis (also sometimes called the Forgetting-Reconstruction Hypothesis ), it is proposed that random practice forces a performer to abandon (or forget) their action plan (GMP) with each new trial, because they are likely moving to executing a ...
The action-plan reconstruction explanation, proposed by Lee and Magill (1983, 1985), emphasizes forgetting and the subsequent reconstruction of a movement plan. In this hypothesis, random practice causes an increase in information processing activities for a given response because the intervening practice causes the learner to forget previously ...
Lin et al. (2008) confirmed the elaborative-processing hypothesis, as in their study, M1 inhibition during the intertrial intervals of the acquisition phase of blocked practice did not generate sufficient forgetting and action plan reconstruction to produce the same level of learning as in random practice. While the performance levels of those ...
In two of the three experiments that are reported subjects were trained in either a blocked or a random practice schedule and allowed to self-select the amount of time they used to plan an upcoming movement. According to the reconstruction hypothesis (Lee & Magill, 1985) random practice participants engage greater movement planning processes (i ...
147 support for this hypothesis would be expected in the perceptual-cognitive skills domain 148 (Broadbent et al., 2015a; Memmert et al., 2009). In contrast, the definition for the action plan 149 reconstruction hypothesis states that for an upcoming task in random practice 'a person must
reconstructive hypothesis (or action plan reconstruction hy-pothesis) posits that interleaving is beneficial because it re-quires each task set to be frequently retrieved, while during blocked practice the task set remains in working memory for theentiretaskduration(Lee&Magill,1983;Leeetal.,1985).
The action-plan reconstruction hypothesis Several authors (Lee & Magill, 1983; Lee, Magill, & Weeks, 1985) have tried to account for the CIE by calling on a 'r econstruction' mechanism that takes
The forgetting-reconstructive hypothesis (or action plan reconstruction hypothesis) posits that interleaving is beneficial because it requires each task set to be frequently retrieved, while during blocked practice the task set remains in working memory for the entire task duration (Lee & Magill, 1983; Lee et al., 1985).
Memmert et al., 2009). In contrast, the definition for the action plan reconstruction hypothesis states that for an upcoming task in random practice "a person must retrieve the appropriate motor program representing that action and then add the pa-This document is copyrighted by the American Psychological Association or one of its allied ...
On the other hand, if reconstruction of the action plan benefits learning, as suggested by the forgetting-recon- struction hypothesis, adding perturbation (by TMS) during
According to the "forgetting-reconstruction" hypothesis of the CI effect, short-term forgetting between successive presentations of the same task during random training requires the learner to "reconstruct the action plan at each presentation," resulting in stronger memory representations (Lee and Magill 1983; Lee et al. 1985). Recent ...
According to this explanation (referred to as the action plan reconstruction hypothesis), random learners forget the movement solution of each task as they process the task requirements of the other. On switching back to the initial task, they are forced to undergo a demanding reconstructive process to replan the way in which they perform the task.
Two notable explanations for this phenomenon include the elaborate processing hypothesis (Shea & Morgan, 1979) and the action-plan reconstruction hypothesis (Lee & Magill, 1983; Lee, Magill, & Weeks, 1985). The elaborate processing hypothesis postulates that stronger memory representations are achieved by comparing differences between tasks (or ...
By mixing up tasks during practice the athlete has to plan, forget and re-plan each time, this is not required when carrying out the same task over and over again (constant-blocked practice). This theory is called the forgetting-reconstruction hypothesis. A similar, but slightly differing theory is called the elaboration hypothesis. Whereby ...
The elaboration distinctiveness hypothesis and the forgetting reconstruction hypothesis have been investigated and offered as an explanation to the CI effects. ... differences in acquisition are reduced. They demonstrated that action plan reconstruction can take place before and after the onset of a movement when the preparation time is limited
The forgetting reconstruction hypothesis (Lee & Magill, 1985) postulates that high CI demands more efforts from learners to develop new plans for accomplishing tasks successfully, as the action plan for the next task would have been forgotten. More specifically, in blocked practice, an action plan is developed by the learner to perform a task ...
Alternatively, the 'forgetting-reconstruction hypothesis' [12][13][14] centers on the idea that random practice results in forgetting between repetitions of the distinct strategies required for ...
These results support the elaborative-processing hypothesis by showing that perturbing information processing, evoked by Random Practice, deteriorates the learning benefit. Unlike the prediction of the forgetting-reconstruction hypothesis, adding perturbation during Blocked Practice did not significantly enhance motor learning.
The forgetting-reconstructive hypothesis (or action plan reconstruction hypothesis) posits that interleaving is beneficial because it requires each task set to be frequently retrieved, while during blocked practice the task set remains in working memory for the entire task duration (Lee & Magill, 1983; Lee et al., 1985).
hypothesis: [noun] an assumption or concession made for the sake of argument. an interpretation of a practical situation or condition taken as the ground for action.